 Sarah L. Hale
Sarah L. Hale John L. Koprowski
John L. Koprowski Steven R. Archer
Steven R. Archer
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 17 July 2020
Sec. Biogeography and Macroecology
Volume 8 - 2020 | https://doi.org/10.3389/fevo.2020.00233
This article is part of the Research Topic Ecological, Behavioral and Genomic Consequences in the Rodent Family Sciuridae: Why Are Squirrels So Diverse? View all 14 articles
Tree and shrub proliferation has been widespread in grasslands worldwide, and has altered ecosystem function and wildlife habitat. Several causes have been proposed for the woody plant encroachment phenomenon. The widespread eradication of a native keystone herbivore in North American grasslands, the prairie dog (Cynomys spp.), is one potential contributing factor that has received relatively little attention. We hypothesized prairie dogs would have historically suppressed woody plants by creating “browse traps” through their systematic clipping of vegetation. We tested this hypothesis by conducting surveys and experimentally manipulating shrub accessibility via exclosures and artificial saplings on and around recently reestablished black-tailed prairie dog (Cynomys ludovicianus) colonies in southeastern Arizona, United States. Shrubs were common on the nascent colonies (mean ± SE = 132 ± 32.7 plants ha–1), but at substantially reduced densities compared to off colonies (305 ± 94.9 plants ha–1). Among branches placed on colonies to simulate “saplings” 89% were damaged within 3 days of “planting,” whereas those placed off colonies were virtually untouched. This was true for both a deciduous, N2-fixing shrub (velvet mesquite, Prosopis velutina) and an evergreen non-N2-fixing shrub (creosote bush, Larrea tridentata). Prairie dogs on newly established colonies did not extirpate woody plants over the time-frame of our study, but reduced their abundance and suppressed their growth, which would ostensibly prevent them from achieving dominance. Implications for extending the longevity of widely practiced “brush management” grassland restoration treatments are discussed in the context of perceptions of prairie dogs as rangeland pests. Prairie dogs represent an enigma in keystone conservation. Whereas the reintroduction of large or charismatic keystone species [e.g., sea otters (Enhydra lutris)] are conducted to restore critical ecological function, reintroductions of other keystone species, such as gray wolves (Canis lupus) and in our case, prairie dogs, are highly controversial. Our findings suggest reintroductions of this negatively perceived small herbivore could function as a tool to locally suppress woody plant proliferation that is widely regarded as an impediment to livestock production. Accordingly, prairie dogs could promote restoration efforts to re-establish and maintain habitat for grassland endemics while promoting biological diversity and other ecosystem services.
Woody plant encroachment, the proliferation of trees and shrubs within grasslands and savannas, is a phenomenon occurring in arid and semiarid grasslands and savannas worldwide. This proliferation may involve increases in the density, stature and cover of native trees and shrubs within their historic ranges or the introduction of non-native woody plants (Archer et al., 2017). Shifts from grass to woody plant dominance affects a variety of ecological processes and the allocation of numerous ecosystem goods, and provisioning (e.g., forage production), supporting (e.g., primary production, biodiversity), and regulating (e.g., carbon sequestration) services (Eldridge et al., 2011; Archer and Predick, 2014). Increases in woody cover also substantially alters wildlife habitat, favoring some species and adversely affecting others (Fulbright et al., 2018). Increases in atmospheric CO2 and changes in climate and fire regimes have been proposed as causal factors; however, the direct and indirect effects of intensification of livestock grazing over the past century appears to be a primary driver (Archer et al., 2017). Perspectives on shrubs in grasslands vary widely (Archer et al., 2017). Commercial ranching enterprises based on cattle have traditionally viewed shrub proliferation as an impediment to livestock production; pastoralists raising goats, camels or other domesticated browsers see shrubs as a potentially important forage resource and may look favorably on increases in their abundance; conservation biologists have biodiversity concerns that shrub proliferation will threaten grassland and savanna ecosystem types and the plants and animals endemic to them (Archer et al., 2017). These contrasting perspectives pose challenges for managing lands for multiple uses.
Coincident with the intensification of livestock grazing in North America was the widespread eradication of the prairie dog (Cynomys spp.), a rodent traditionally viewed as an impediment to livestock production (Vermeire et al., 2004; Derner et al., 2006; Detling, 2006). Although prairie dogs have been recognized as contributing to the maintenance of grassland ecosystems (Hoogland, 1995; Kotliar et al., 1999), the consequences of their removal have received relatively little attention in the context of regulating grass-woody plant interactions. In Africa, herbivores of varying sizes mediate woody plant abundance through browsing activities [e.g., elephants (Loxodonta spp.), dik-diks (Madoqua spp.), impala (Aepyceros melampus); Sankaran et al., 2013]. Systematic browsing maintains woody vegetation at a small stature, thereby creating a “browse trap” that also contributes to a “fire trap” (Bond and Keeley, 2005; Staver et al., 2009; Sankaran et al., 2013). For the purpose of this manuscript, “browse trap” and “fire trap” refer to the maintenance of woody vegetation at small stature by herbivores and frequent, low-intensity wildfires, respectively. Woody plants caught in a fire or browse trap are unable to achieve dominance until the suppressing pressure is removed. We suggest that prairie dogs in North America historically prevented woody plant recruitment or created and maintained browse traps that suppressed shrubs and trees. In the former case, prairie dog eradication would have opened the door for woody plant establishment. In the latter case, prairie dog removal would have released established, but diminutive, woody plants from the browse trap. In both cases, removal of the suppressing pressure, i.e., prairie dogs, would have permitted woody plants to reach adult stature in grasslands throughout the prairie dog’s former range.
Prairie dogs, once widely distributed across the North American west (Proctor et al., 2006), were deemed a rangeland pest in competition with livestock, leading to widespread and highly successful eradication efforts beginning in the early 1900s (Andelt, 2006; Forrest and Luchsinger, 2006; Reeve and Vosburgh, 2006). Historic population sizes are hard to pin-point (Vermeire et al., 2004), but at the most conservative estimate, active eradication, habitat loss, and sylvatic plague (Yersinia pestis; Luce et al., 2006) appear to have reduced prairie dog populations to 2% of historic numbers (Whicker and Detling, 1988; Miller et al., 1994), and range occupancy to ≤1% (Luce et al., 2006). Prairie dogs are grazers, but systematically gnaw, clip, and girdle other vegetation to maintain plants on their colonies at small stature (<30 cm; King, 1955; Hoogland, 1995; Weltzin et al., 1997) and thereby maximize predator detection. Accordingly, their removal would have removed an impediment to woody plant establishment and growth.
Woody plant proliferation in grasslands has led to the implementation of “brush management” practices involving heavy equipment, herbicides, and/or prescribed fire to reduce the cover of trees and shrubs (Hamilton et al., 2004; Collins et al., 2015). Such techniques are typically expensive with short-lived effects, and hence are seldom cost-effective (Archer et al., 2011). We sought to investigate whether the black-tailed prairie dog (Cynomys ludovicianus; hereafter “prairie dog”) might constitute a natural, potentially long-term, sustainable “biocontrol” solution for deterring woody plant encroachment and proliferation in grasslands. We hypothesized that reintroduced prairie dogs would suppress woody plant growth on their colonies, and create a “browse trap” through their systematic felling of vegetation. If our hypothesis was supported, we predicted that woody plants on newly established prairie dog colonies would be less abundant than those off-colony, that shrubs occurring outside of exclosures on prairie dog colonies would have slower growth and higher mortality rates than protected shrubs, and that disturbance to introduced artificial “simulated saplings” would be greater on colonies than off colonies.
From a broader perspective, our study was also intended to determine whether the reintroduction of a keystone herbivore following a prolonged absence could restore the browse trap that historically excluded woody plants or prevented them from expressing dominance in grassland systems. Studies involving keystone species typically seek to assess ecosystem effects while the species of interest is present or absent from the landscape (e.g., Reisewitz et al., 2006); however, studies assessing the ecosystem-level effects of keystone species reintroductions are more rare (Hale and Koprowski, 2018). A study in northwestern Mexico investigated independent and interactive effects of existing prairie dogs and cattle on woody encroachment in an elegant field experiment (Ponce-Guevara et al., 2016), wherein plots received one of four treatments: prairie dogs only, cattle only, prairie dogs + cattle, and neither prairie dogs nor cattle. Results indicated that prairie dogs played an important role in suppressing woody encroachment, and that their role was amplified in the presence of cattle. The study we report here similarly assesses the effects of black-tailed prairie dogs on shrubs in an arid grassland, but differs in that we had the unique opportunity to ascertain whether a reintroduced population of prairie dogs could influence shrub encroachment. Our study is among the first to investigate consequences of the reintroduction of a keystone species, and the extent to which that reintroduction might be justified as part of an integrated, comprehensive plan to more effectively address ecosystem management challenges in grasslands.
Las Cienegas National Conservation Area (hereafter Las Cienegas) is a 17,000 ha working cattle ranch managed by the Bureau of Land Management, and is located 72 km southeast of Tucson, AZ, United States in Santa Cruz County. The study area is a representative of the natural regional grasslands occurring across central and southern Arizona, southwestern New Mexico, and northern Mexico (Gori and Enquist, 2003; Gori and Schussman, 2005; Bodner and Simms, 2008). Mean annual precipitation is 405 mm, mostly falling during the summer monsoon months (July–September); mean annual temperature is 15.7°C (Bodner and Robles, 2017). During our study (2013–2015) annual precipitation and temperature (±SE) averaged 350 ± 47 mm and 16.2 ± 0.2°C (Empire Remote Area Weather Station; QEMA3)1. Grasslands at Las Cienegas are located between 1300 and 1500 m in elevation, and soils are primarily gravely, sandy, and clay loam (Bodner and Robles, 2017).
Semi-desert grasslands in Arizona support a variety of herbivores, including peccaries (Pecari tajacu), mule deer (Odocoileus hemionus), white-tailed deer (Odocoileus virginianus), pronghorn (Antilocapra americana), and historically, supported black-tailed prairie dogs, which were extirpated by 1960 (Bock and Bock, 2000; Underwood and Van Pelt, 2000). The Arizona Game and Fish Department began reintroducing black-tailed prairie dogs at Las Cienegas in 2008 using animals obtained from populations in New Mexico, United States, and Sonora, Mexico. We conducted our experiments on four of these colonies.
Colonies were situated on sites with similar soils (fine, deep, well-drained), slopes (0–15%), and elevations (1367–1412 m). One colony was initiated per year starting in 2008 and ending in 2011. The Arizona Game and Fish Department prepared sites via mechanical removal of large (>∼1 m height) shrubs, mowing, and installation of 25 artificial burrows within a 4 ha area (see Hale et al., 2013 for details). Managers re-mowed colonies in 2011 to facilitate prairie dog establishment.
Throughout our study, three of the four colonies remained occupied by prairie dogs; however, one colony (hereafter referred to as the “control site”) failed and was unoccupied during 2014 and 2015. The Arizona Game and Fish Department initially prepared this site in the same manner as the other colonies, but the colony lacked prairie dogs throughout the majority of our study period (<10 prairie dogs present in May 2013; the site was vacant by October 2013). We only used the control site for our simulated sapling experiment.
We surveyed woody plants on each occupied colony in May of 2013 and 2014. We conducted eight belt (5 m width) transect surveys, wherein transects originated at the center of each colony and extended to and slightly beyond the colony boundary. We considered colony boundaries to be where the junction of short, prairie dog- defoliated grass and burrows met tall, unclipped grass and an absence of burrows. Off-colony transects extended up to 100 m beyond the colony perimeter for four transects at colonies A and B, and six transects at colony C. Some off-colony transects did not extend to the full 100 m due to logistical or topographic constraints (e.g., one transect was cut short by a deep, wide wash). We counted shrubs with stems originating within the transect belts, and noted the presence and degree of disturbance in the form of clipping, chewing, or girdling of their stems on a scale of 0–3 (0 = no disturbance, 1 = few stems disturbed, 2 = many stems disturbed, 3 = majority of stems disturbed). We considered colony to be the experimental unit (n = 6; two surveys at three colonies).
Among the woody plants encountered in our 2013 survey, we tagged plants of a random subset of the two most abundant species (velvet mesquite, Prosopis velutina and catclaw acacia, Acacia greggii; n = 40) for monitoring on each occupied colony (typically n = 20 on-colony and 20 off-colony). We installed exclosures around half of the plants at each on-off colony location (n = 10), with the other half remaining unprotected. We used garden fencing (mesh size 5.1 cm × 7.6 cm) to exclude prairie dogs and larger mammals (e.g., lagomorphs) for half of the exclosures (n = 5), and hardware cloth (mesh size 0.6 cm × 0.6 cm) for the remainder of the exclosures (n = 5) to exclude prairie dogs and smaller mammals [e.g., northern pygmy mice (Baiomys taylori), pocket mice (Chaetodipus spp., Perognathus spp.), kangaroo rats (Dipodomys spp.), woodrats (Neotoma spp.), grasshopper mice (Onychomys spp.), deer mice (Peromyscus spp.), harvest mice (Reithrodontomys spp.), and cotton rats (Sigmodon spp.)]. We left exclosures in place for ∼3 months (mid-May through August) in 2013 and 2014 and for ∼5 months (mid-May through late October) in 2015. We measured the canopy diameter of plants along their longest axis and perpendicular to their longest axis at the time of exclosure installation and again at the time of exclosure removal. We standardized change in shrub canopy area (CA), (m2, computed as an ellipse) to a monthly basis (30 days). We followed the same plants each year; if a plant measured on the previous year had died, we made note of it and replaced it with the nearest live individual of the same species.
We removed foliated branches [average (±SE) length = 47 ± 0.6 cm; diameter = 1.1 ± 0.02 cm] from mature shrubs in nearby off-colony areas, and inserted them ∼10 cm into the ground along four transects that originated at the center of each colony and extended 100 m beyond the colony boundary in randomized directions. We also installed transects of “simulated saplings” on the unoccupied control site that had been prepared in the same manner as the occupied colonies (see section “Study Site”).
We hypothesized that prairie dog responses to artificial “simulated saplings” would indicate how they would react to the appearance of new woody plants. We spaced saplings (n = 10) evenly along transect segments on the colonies (the spacing distance varied for each transect segment, depending on its absolute length), and at 10 m intervals along the off-colony transect segments (n = 10). We considered three simulated saplings on each transect to be “periphery” saplings: the sapling installed on the colony boundary, and the closest sapling inside and outside of the colony boundary (Supplementary Figure S1). We also sought to ascertain whether prairie dogs would respond to familiar vs. novel shrubs in the same manner, so we used two woody species: velvet mesquite (P. velutina; hereafter “mesquite”), a native shrub common on and around the prairie dog colonies, and creosote bush (Larrea tridentata), also a native shrub, but one that did not occur on or near the colonies on the Las Cienegas. Both species are abundant across the Sonoran Desert and on landscapes within the study area. Mesquite is a thorny, N2-fixing deciduous shrub (Fabaceae), whereas creosote bush is thorn-free, non-N2-fixing evergreen shrub (Zygophyllaceae) that dominates many landscapes in each of the three North American hot deserts.
We measured saplings for height (cm) and recorded the number of secondary stems arising from the primary stem (typically several for mesquite and none for creosote bush). After 3 days of exposure to prairie dogs, we re-measured saplings and recorded the extent of damage (i.e., chewing, clipping, or girdling). We quantified damage as the proportion of stems disturbed on mesquite saplings and the relative proportion of height change of creosote bush saplings (since there were rarely secondary stems present). We conducted the experiment with mesquite in May 2014, and repeated it in late July/early August 2014. We conducted the experiment with creosote bush saplings in early September 2015, and repeated it in late September 2015. We generated different sets of random transects for each trial, so considered temporal replicates independent of one another for statistical analysis. For our 2014 experiment, we used branches of various heights (11–156 cm) and basal diameters (2.2–24.5 mm) to ascertain if prairie dog treatment of small, less visible plants would differ from that of larger more prominent plants.
We performed statistical analyses in JMP (1989-2007). We assessed shrub density on and off colonies using a Student’s t-test, with location (on-colony, off-colony) as the explanatory variable. We removed outliers (points > ± 2 SD’s from the mean; n = 7 of 320 shrub canopy area data points) from analysis based on examination of histograms and residual plots. We log transformed changes in shrub canopy area (m2/month) in exclosure experiments to normalize the data and assessed relationships using a full factorial two-way ANOVA with exclosure mesh size (prairie dogs and larger mammals excluded, or all mammals excluded) and location (on-colony, off-colony) as main effects. Additionally, we assessed change in shrub canopy area using a four-way ANOVA with location (on-colony, off colony), exclosure presence (yes, no), colony (A, B, or C), and year (2013, 2014, 2015) as main effects.
To assess the simulated sapling experiment, we performed a full factorial three-way ANOVA with trial number (first or second), species (mesquite or creosote bush), and location (on-colony, colony periphery, off-colony, on-control, control periphery, off-control) as independent variables, and proportion of total plants disturbed or degree of disturbance as the dependent variables. We considered colony to be the experimental unit (n = 16; four temporal replicates per colony).
When significant differences were indicated by ANOVA (α < 0.05), we used Tukey’s honest significant difference (hsd) tests (where statistical significance was set at 0.05 divided by the total number of comparisons in each test) to evaluate differences among means. In cases where no interactions occurred between effects having only two levels, we performed Student’s t-tests on each effect with α < 0.05.
Mean (±SE) shrub density (plants ha–1) was greater off of colonies (305 ± 94.9) than on colonies (132 ± 32.7; P = 0.07; Figure 1). The frequency of disturbance was greatest for shrubs on colonies [85% (n = 329 plants)] compared to only 9% (n = 279) in off-colony locations.
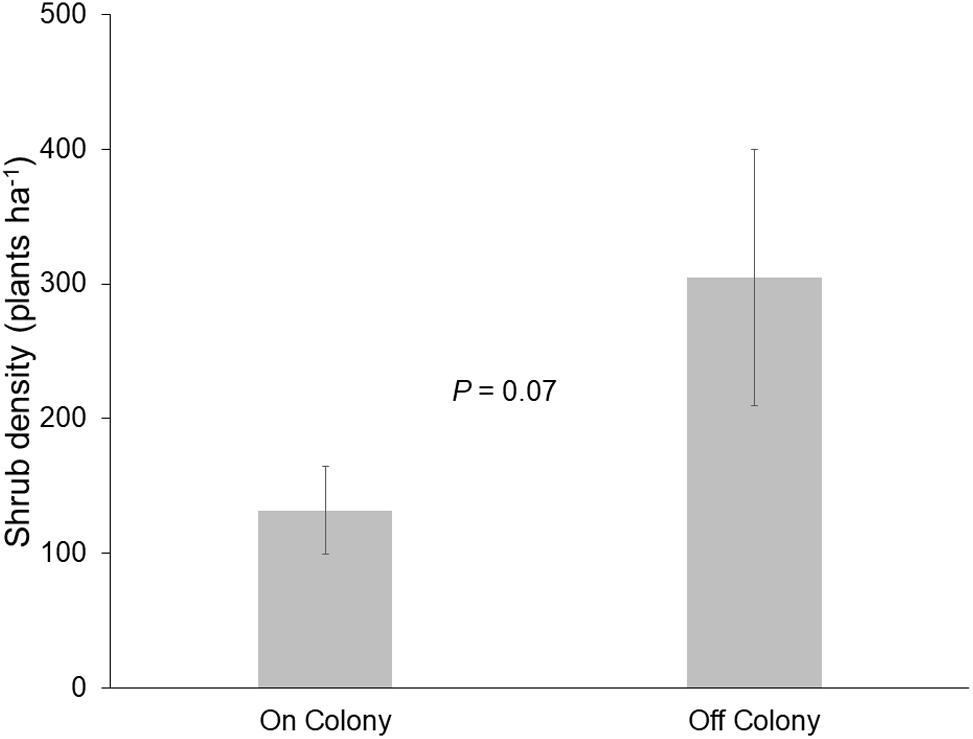
Figure 1. Mean (±SE) density of shrubs (Prosopis velutina and Acacia greggii) on and off of black-tailed prairie dog (Cynomys ludovicianus) colonies (n = 6; two surveys at three colonies).
Shrub mortality during our study was low: only three of the 116 tagged mesquite plants died. Survival of mesquite was comparable on (95 ± 2.0%) and off of colonies (100 ± 2.0%; P = 0.10), and inside (95 ± 2.0%) and outside of exclosures (100 ± 2.0%; P = 0.10).
Changes in the canopy area of individual shrubs (m2/month) were statistically comparable in exclosures of differing mesh size (F1,159 = 0.01, P = 0.918; Table 1) so we pooled these data for subsequent analysis. Additionally, changes in shrub canopy area were statistically comparable among colonies (F = 0.412, P = 0.663; Table 1), so we also pooled these data for subsequent analysis. We examined data to determine if there might be a size threshold above which shrubs might be more consistently damaged by prairie dogs, but we found no indication of such.
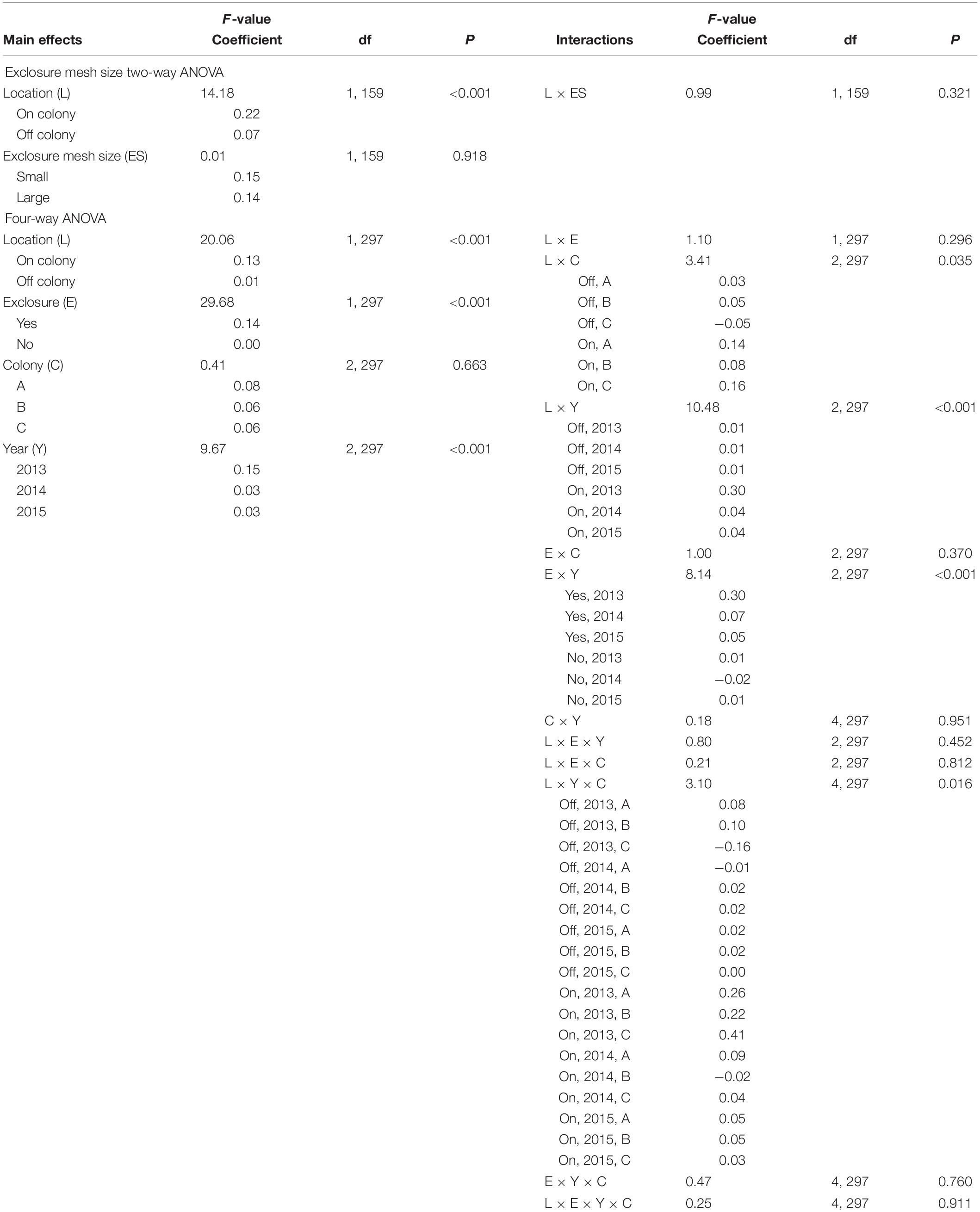
Table 1. Summary of ANOVA outcomes for exclosure experiments and coefficients for main effects and significant interactions.
Change in canopy area was affected by location (F2,297 = 20.06, P < 0.001; Table 1), year (F2,297 = 9.67, P < 0.001; Table 1), exclosure presence (F2,297 = 29.68, P < 0.001; Table 1), and the interactive effects of location × year (F2,297 = 10.48, P < 0.001; Table 1) and exclosure presence × year (F2,297 = 8.14, P < 0.001; Table 1). On average, (i) canopies of shrubs protected by exclosures expanded more than those of unprotected shrubs (P < 0.001), (ii) levels of canopy expansion of shrubs on colonies (0.14 ± 0.03 m2/month) was greater than that of shrubs off colonies (0.00 ± 0.02 m2/month; P < 0.001), and (iii) canopies of shrubs expanded more in 2013 (0.15 ± 0.02 m2/month) than in 2014 (0.03 ± 0.02 m2/month; P < 0.001) or 2015 (0.03 ± 0.02 m2/month; P < 0.01). Interaction effects were largely driven by year, with shrubs on colonies in 2013 expanding significantly more than all other groups (Table 1), and shrubs inside exclosures in 2013 expanding more than all other groups (Table 1).
Size of simulated saplings did not affect whether or not they were disturbed [X2 (df = 3, n = 638) = 1.309, P = 0.73], so we pooled size classes for subsequent analysis. Colony had no effect on the proportion of saplings disturbed or degree of disturbance, so we pooled these data across occupied colonies as well. Neither trial number (first or second; F1,47 = 0.17, P = 0.688) nor shrub species (creosote or mesquite; F1,47 = 0.19, P = 0.671) influenced the proportion of saplings disturbed, but location (on-colony, colony periphery, or off-colony, on-control, control periphery, off-control) did (F1,47 = 61.63, P < 0.001). The proportion of saplings disturbed was greatest on colonies (89 ± 4.0%; see Figure 2 for illustration of disturbance), followed by colony peripheries (39 ± 4.0%; Tukey’s hsd, P < 0.001); disturbances off-colony were essentially nil (0–2%; Figure 3A). Expressed as a spatial continuum, the proportion of saplings disturbed was consistently high (74–100%) among on-colony transect positions, decreasing sharply at the colony periphery locations, and was consistently low to nil for off-colony positions and the control site (Figure 4).

Figure 2. Simulated velvet mesquite (Prosopis velutina) sapling (approximately 30 cm in height) upon installation on an active black-tailed prairie dog (Cynomys ludovicianus) colony (left) vs. after 3 days of exposure to black-tailed prairie dogs (right).
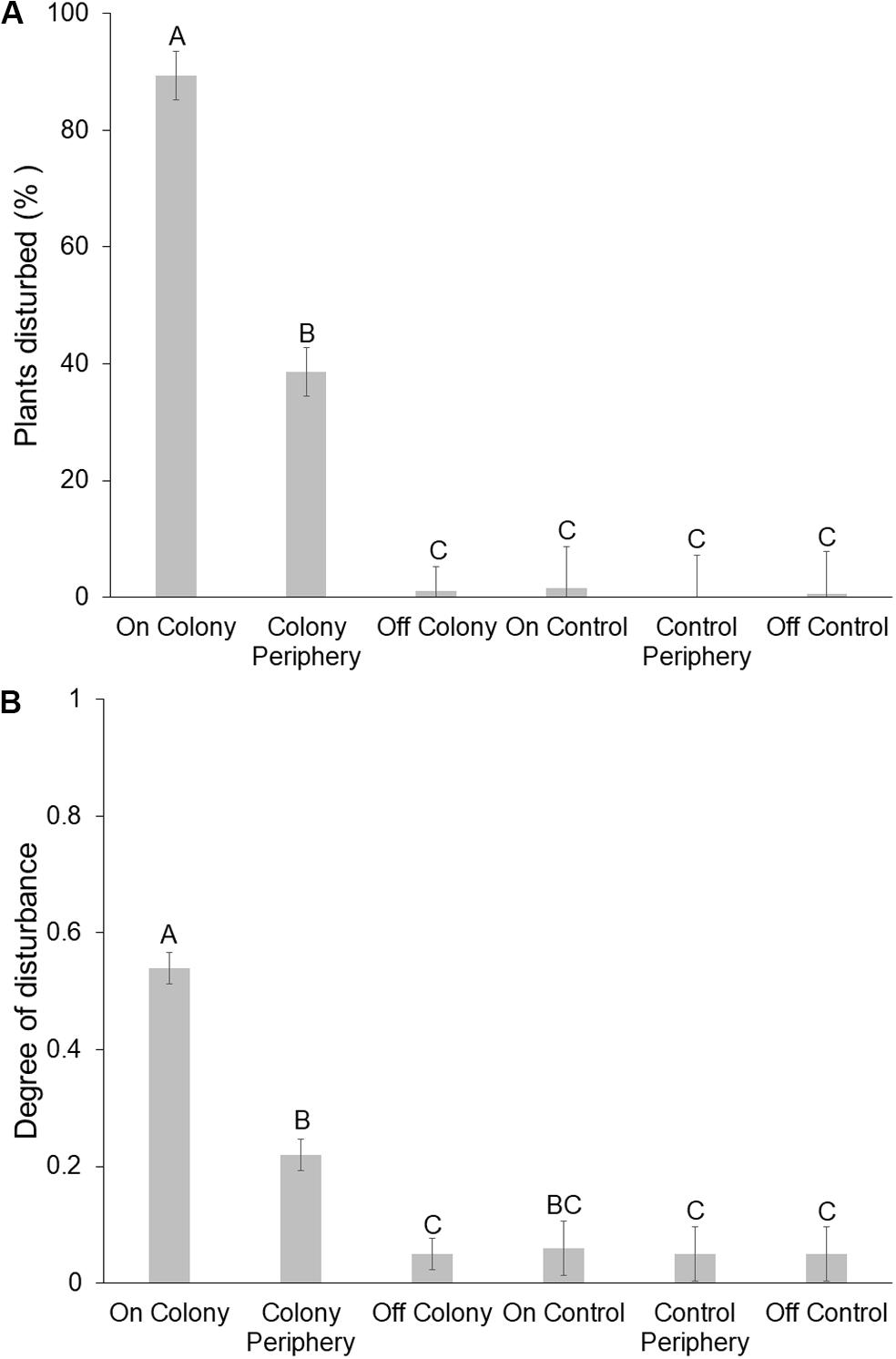
Figure 3. Mean (±SE) percentage of “simulated” Prosopis velutina and Larrea tridentata saplings disturbed by black-tailed prairie dogs (Cynomys ludovicianus) (A) and scalar (0 = none; 1 = max) assessments of the degree of disturbance (B) on, on peripheries, and off of active colonies and a control site (n = 16; see section “Materials and Methods”). Different letters above bars indicate significant (P < 0.05) differences between means within a panel.
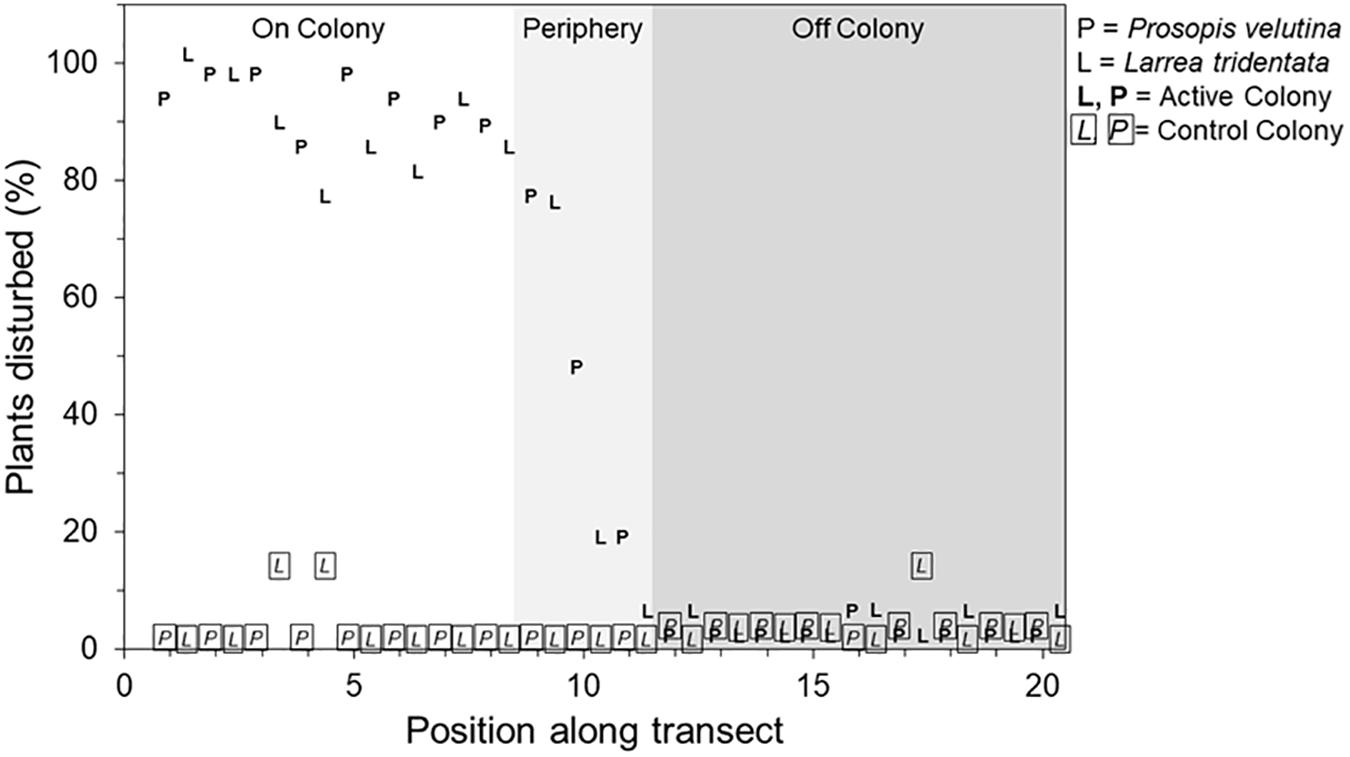
Figure 4. Mean percentage of simulated Prosopis velutina (denoted “P”) and Larrea tridentata “saplings” (denoted “L”) disturbed along transects originating at the centers (position = 0) of active black-tailed prairie dog (Cynomys ludovicianus) colonies (bold font) and a control colony (italic font, boxed). Transects varied in length, depending upon the size of the colony, so positions are relativized such that 1–8 represent locations on colony, 9–11 represent locations on the colony periphery, and 12–20 denote the off colony locations (see Supplementary Figure S1).
Similarly to the proportion of plants disturbed, neither trial number (first or second; F1,47 = 1.43, P = 0.243) nor shrub species (creosote or mesquite; F1,47 = 2.27, P = 0.145) influenced the degree of sapling disturbance. Conversely, location (on-colony, colony periphery, or off-colony, on-control, control periphery, off-control) was significant (F1,47 = 44.35, P < 0.001). Degree of sapling disturbance was greatest on colonies (0.54 ± 0.03), intermediate on colony peripheries (0.22 ± 0.03), and least off of colonies (0.05 ± 0.03) and on control sites (on: 0.06 ± 0.05, periphery: 0.05 ± 0.05, off: 0.05 ± 0.05; Figure 3B).
Woody plant encroachment is a threat to grasslands and their endemic plants and animals (Archer et al., 2017). Chemical, mechanical and pyric approaches to reducing woody plant abundance and restoring shrub-encroached grasslands have been widely practiced (collectively known as “brush management”), but are typically expensive and their effects are short-lived (Archer and Predick, 2014). Our results suggest that the reintroduction of prairie dogs subsequent to “brush management” could prolong the effectiveness of those treatments while concurrently resuscitating ecosystem processes that would sustain long-term success and promote biodiversity. In this respect, prairie dog re-introductions represent another example of how “rewilding” may revive ecosystem structure, function, and health in a cost-effective manner (e.g., Guyton et al., 2020).
Our exclosure experiments indicated that herbivory was reducing shrub growth both on and off the colony, but more so off-colony than on-colony. One hypothesis to account for this counter-intuitive result would be that above- vs. belowground allocation patterns differed for shrubs on- and off-colony, wherein on-colony shrubs were preferentially allocating more resources to aboveground growth. Another hypothesis would be that growth of protected on-colony shrubs was enhanced by reductions in competition resulting from the felling of surrounding plants by prairie dogs, while growth of protected off-colony shrubs was muted or suppressed owing to the relatively higher levels of competition from surrounding plants. In any case, our observations, surveys, and sapling experiments collectively suggest that benefits gained by potential release from competition or increased allocation to aboveground growth were more than offset by high levels of damage to on-colony shrubs. We directly observed prairie dogs gnawing and clipping shrubs (including those that escaped the broad-scale mechanical treatment and those that re-sprouted following mechanical treatment) and our simulated saplings, consistent with their well-documented habit of systematically felling plants to maintain vegetation in a short-statured state to aid in visual detection of predators (King, 1955; see Figure 5.18 in Hoogland, 1995). Small-statured shrubs that remained on colonies after the mechanical removal of adult shrubs were disturbed more frequently than those located on colony peripheries and off of colonies, as substantiated by observational results (surveys), and experienced the greatest degree of disturbance as indicated by our simulated sapling experiments. Prairie dogs were thus able to substantively mitigate shrub abundance and stature on these nascent colonies. The introduced prairie dogs did not eliminate the small shrubs remaining on their colonies over the time-frame of this study, but the density of shrubs on colonies was 40% that of off-colony sites (Figure 1). This suggests that prairie dogs may reduce recruitment and, over longer time-scales, induce shrub mortality on their colonies.
Prosopis velutina and A. greggii, the two main shrubs on our site, have the ability to vegetatively regenerate from basal meristems following removal of aboveground biomass (Glendening and Paulsen, 1955; Cross and Wiedemann, 1997). Accordingly, prairie dogs on these nascent colonies did not affect shrub survival, demonstrating that shrubs at this site are capable of persisting even under heavy levels of herbivory. This is reminiscent of the “browse trap” phenomenon. We would expect that the loss of prairie dogs from this site, which had been mechanically cleared of adult woody plants, would release small shrubs or those recruiting from the seed bank from the browse trap. These expectations are consistent with those from studies on well-established prairie dog colonies (Weltzin et al., 1997; Ceballos et al., 2010; Ponce-Guevara et al., 2016). Such a release would allow shrubs to re-gain dominance and thus neutralize the “brush management” effort to restore this former grassland. The fact that simulated saplings placed on a recently failed prairie dog colony received no damage (Figures 3, 4), despite their being highly conspicuous to lagomorphs and other rodents, suggests that prairie dogs, not other small mammals, were regulating shrubs within colonies. This interpretation is consistent with another study showing that small rodents on prairie dog colonies did not influence rates of shrub seed disappearance (Weltzin et al., 1997).
There was little evidence that large herbivores contributed to a browse trap for shrubs in this region. Native (e.g., American bison, Bison bison) and non-native ungulates, such as domestic cattle (Bos taurus) have diets that consist primarily of grasses and forbs (Meagher, 1986; Beck and Peek, 2005) so their preferential foraging on prairie dog colonies (e.g., Coppock et al., 1983) would not be likely to constrain the development of communities dominated by woody plants prior or subsequent to prairie dog removal. This would be particularly so for highly unpalatable shrubs such as mesquite and creosote bush (Campbell and Taylor, 2006; Brock et al., 2014; Bovey, 2016). Pronghorn (Antilocapra spp.) are also known to preferentially forage on prairie dog colonies (King, 1955; Krueger, 1986; Hoogland, 1995), but given the abundance of shrubs in other portions of their range [e.g., sagebrush (Artemesisia spp.) in the Intermountain West and northern Great Plains] it does not seem likely they exert sufficient browsing pressure to keep shrubs in check. The browse trap in this system thus appears to center around prairie dogs and their activities, which could, in turn, be enhanced by cattle grazing (Ponce-Guevara et al., 2016).
A coincidence between the elimination of small native grazers and woody encroachment is not restricted to North America. In Australia, native granivorous rodents may have historically limited shrub recruitment following fire events, but rodent populations have declined due to pastoral activity, and their declines have coincided with shrub encroachment (Gordon and Letnic, 2019). Plains vizcacha (Lagostomus maximus) in Argentina and burrowing bettong (Bettongia lesueur) in Australia exhibit similarities to prairie dogs not only in their ecology and behavior, but also in their pest status, which has led to elimination from parts of their range (Jackson et al., 1996; Noble et al., 2007). Likewise, plateau pikas (Ochotona curzoniae) in China play a keystone role analogous to that of prairie dogs, but have also been subjected to eradication efforts (Smith and Foggin, 1999; Delibes-Mateos et al., 2011; Fahong et al., 2012). As with the elimination of prairie dogs in North America, elimination of these perceived small mammalian pests has had the unintended consequence of promoting woody plant proliferation (Brandt et al., 2013; González-Roglich et al., 2015). Such trade-offs should be considered in rangeland management and conservation plans where the maintenance of grasslands and the organisms endemic to them are a priority.
Woody plant proliferation in grasslands has prompted the implementation of “brush management” practices aimed at reducing shrub cover to enhance livestock production (Hamilton et al., 2004) and restore habitat for grassland wildlife (Fulbright et al., 2018). These practices often involve herbicides, heavy equipment (e.g., cutting, shredding, or chaining), and prescribed burning alone or in combination (Collins et al., 2015). Such endeavors are costly, short-lived, and require repeated application (Archer et al., 2011). Prairie dogs rely on short vegetation for visual detection of predators (Hoogland, 1995; King, 1955), so are unlikely to colonize areas with dense woody vegetation and would not be likely to persist if introduced into a landscape with well-developed woody cover (Milne-Laux and Sweitzer, 2006). However, our study suggests that reintroductions of prairie dogs following brush management can succeed and could extend the shrub treatment longevity and reduce or perhaps eliminate the need for follow-up treatments, while simultaneously promoting the restoration of grassland habitats and the plants and animals endemic to them (Whicker and Detling, 1988; Kotliar et al., 1999, 2006; Ceballos et al., 2010).
Our sapling experiments suggest that prairie dogs are not limited by previous experience with woody species, as the Las Cienegas prairie dogs felled both a species they encountered regularly (P. velutina) as well as a species they had never encountered (L. tridentata). This suggests that (i) prairie dogs can restore the browse trap even when source and reintroduction sites differ in woody species composition, and (ii) prairie dog reintroduction within areas of their former range could simultaneously function as a natural, cost-effective, and sustainable option for limiting woody plant encroachment in present-day grasslands and for restoring grassland structure, function and biodiversity where shrub encroachment has occurred. These positive facets of prairie dogs may compensate for or more than off-set the traditional negative perspectives associated with prairie dog-livestock competition for forage and declines in forage and livestock production (O’Meilia et al., 1982; Derner et al., 2006). A comprehensive assessment of these complexities in livestock-prairie dog interactions and the various trade-offs that emerge would require a landscape-scale accounting of livestock grazing intensity, seasonality, and patterns and prairie dog colony locations, sizes and ages. When prairie dog re-introductions are contemplated, where best to locate them and how colonies might subsequently expand should be given careful consideration.
The conservation and reintroduction of large or charismatic keystone species [e.g., gray wolves (Canis lupus) and sea otters (Enhydra lutris)] can restore critical ecological function to ecosystems (Estes and Palmisano, 1974; Callan et al., 2013). However, similar to the historical perception of wolves, small keystone herbivores are often viewed as pests that are targeted for eradication rather than conservation (Delibes-Mateos et al., 2011). Accordingly, the ecological implications of reintroducing small keystone herbivores has not been widely investigated. We add to this understanding, and highlight the ability of small keystone herbivores to address significant ecological issues, such as woody encroachment, immediately following their reintroduction and in conjunction with other land management practices. Our results indicate the important role(s) that small keystone herbivores can potentially play in maintaining diverse, healthy ecosystems, and the substantial ecological consequences that can result from their removal (Weltzin et al., 1997; Davidson et al., 1999, 2012, 2014; Miller et al., 2000; Kotliar et al., 2006; Brandt et al., 2013; González-Roglich et al., 2015). A better understanding of such species and their reintroduction after extirpation will be important for the restoration and conservation of grassland ecosystems throughout the world.
The datasets generated for this study are available on request to the corresponding author.
The animal study was reviewed and approved by the University of Arizona Institutional Animal Care and Use Committee (protocol 11-251).
SH contributed to the research design, collected data in the field, performed the experiments, analyzed the data, and wrote the manuscript. JK and SA contributed to the research design, data analysis, and provided valuable edits and revisions to the manuscript. All authors contributed to the article and approved the submitted version.
This work was funded by the National Fish and Wildlife Foundation, the Arizona Center for Nature Conservation, the Arizona Game and Fish Department, T & E, Inc., the Bureau of Land Management, and Arizona Agricultural Experiment Station projects ARZT-1360540-H12-199 and ARZT-1390040-M12-205.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Kirsten Fulgham, Paisley and Pandora Hale, Allyson Hawkins, Levi Heffelfinger, Theresa Huckleberry, Max Mazzella, Rachel More-Hla, Rocio Ojeda, Samantha Springs, Michael Taylor, and Kaycie Waters for assisting with field experiments and data collection. We also thank Patricia King (King’s Anvil Ranch) for allowing collection of creosote from her property for simulated sapling experiments and Katherine Predick for assisting with statistical analyses.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.00233/full#supplementary-material
Andelt, W. F. (2006). “Methods and economics of managing prairie dogs,” in Conservation of the Black-Tailed Prairie dog: Saving North America’s Western Grasslands, ed. J. L. Hoogland (Washington, DC: Island Press), 129–138.
Archer, S. R., Andersen, E. M., Predick, K. I., Schwinning, S., Steidl, R. J., and Woods, S. R. (2017). “Woody plant encroachment: causes and consequences,” in Rangeland Systems: Processes, Management and Challenges, ed. D. D. Briske (New York, NY: Springer), 25–84. doi: 10.1007/978-3-319-46709-2_2
Archer, S. R., Davies, K. W., Fullbright, T. E., McDaniel, K. C., Wilcox, B. P., and Predick, K. I. (2011). “Brush management as a rangeland conservation strategy: a critical evaluation,” in Conservation Benefits of Rangeland Practices: Assessment, Recommendations, and Knowledge Gaps, ed. D. D. Briske (Washington, DC: United States Department of Agriculture, Natural Resources Conservation Service), 105–170.
Archer, S. R., and Predick, K. (2014). An ecosystem services perspective on brush management: research priorities for competing land use objectives. J. Ecol. 102, 1394–1407. doi: 10.1111/1365-2745.12314
Beck, J. L., and Peek, J. M. (2005). Diet composition, forage selection, and potential for forage competition among elk, deer, and livestock on aspen-sagebrush summer range. Rangeland Ecol. Manag. 58, 135–147. doi: 10.2111/03-13.1
Bock, C. E., and Bock, J. H. (2000). The View from Bald Hill: Thirty Years in An Arizona Grassland. Berkeley, CA: University of California Press.
Bodner, G. S., and Robles, M. D. (2017). Enduring a decade of drought: patterns and drivers of vegetation change in a semi-arid grassland. J. Arid Environ. 136, 1–14. doi: 10.1016/j.jaridenv.2016.09.002
Bodner, G. S., and Simms, K. (2008). State of the Las Cienegas National Conservation Area. Part 3. Condition and Trend of Riparian Target Species, Vegetation, and Channel Geomorphology. Tucson, AZ: The Nature Conservancy of Arizona, 69.
Bond, W. J., and Keeley, J. E. (2005). Fire as a global ‘herbivore’: the ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 20, 387–394. doi: 10.1016/j.tree.2005.04.025
Brandt, J. S., Haynes, M. A., Kuemmerle, T., Waller, D. M., and Radeloff, V. C. (2013). Regime shift on the roof of the world: alpine meadows converting to shrublands in the southern Himalayas. Biol. Conserv. 158, 116–127. doi: 10.1016/j.biocon.2012.07.026
Brock, J., Brandau, B., Arthun, D., Humphrey, A. L., Dominguez, G., and Jacobs, A. (2014). Long-term results of teburthiuron herbicide treatment on creosote bush (Larrea tridentate) in southeast Arizona, USA. J. Arid Environ. 110, 44–46. doi: 10.1016/j.jaridenv.2014.05.021
Callan, R., Nibbelink, N. P., Rooney, T. P., Wiedenhoeft, J. E., and Wydeven, A. P. (2013). Recolonizing wolves trigger a trophic cascade in Wisconsin (USA). J. Ecol. 101, 837–845. doi: 10.1111/1365-2745.12095
Campbell, E., and Taylor, C. A. (2006). “Targeted grazing to manage weedy brush and trees,” in Targeted Grazing: A Natural Approach to Vegetation Management and Landscape Enhancement, ed. K. Launchbaugh (Centennial, CO: American Sheep Industry Association), 77–88.
Ceballos, G., Davidson, A., List, R., Pacheco, J., Manzano-Fischer, P., and Santos-Barrera, G. (2010). Rapid decline of a grassland system and its ecological and conservation implications. PLoS One 5:e8562. doi: 10.1371/journal.pone.0008562
Collins, C. D. H., Kautz, M. A., Tiller, R., Lohani, S., Ponce-Campos, G., and Hottenstein, J. (2015). Development of an integrated multiplatform approach for assessing brush management conservation efforts in semiarid rangelands. J. Appl. Remote Sens. 9:096057. doi: 10.1117/1.JRS.9.096057
Coppock, D. L., Ellis, J. E., Detling, J. K., and Dyer, M. I. (1983). Plant-herbivore interactions in a North American mixed-grass prairie. II. Responses of bison to modification of vegetation by prairie dogs. Oecologia 56, 10–15. doi: 10.1007/bf00378211
Cross, B. T., and Wiedemann, H. T. (1997). Control of catclaw acacia and mimosa by grubbing. Appl. Eng. Agric. 13, 291–293. doi: 10.13031/2013.21591
Davidson, A. D., Detling, J. K., and Brown, J. H. (2012). Ecological roles and conservation challenges of social, burrowing, herbivorous mammals in the world’s grasslands. Front. Ecol. Environ. 10:477–486. doi: 10.1890/110054
Davidson, A. D., Friggens, M. T., Shoemaker, K. T., Hayes, C. L., Erz, J., and Duran, R. (2014). Population dynamics of reintroduced Gunnison’s prairie dogs in the southern portion of their range. J. Wildlife Manage. 78, 429–439. doi: 10.1002/jwmg.681
Davidson, A. D., Parmenter, R. R., and Gosz, J. R. (1999). Responses of small mammals and vegetation to a reintroduction of Gunnison’s prairie dogs. J. Mammal. 80, 1311–1324. doi: 10.2307/1383181
Delibes-Mateos, M., Smith, A. T., Slobodchikoff, C. N., and Swenson, J. E. (2011). The paradox of keystone species persecuted as pests: a call for the conservation of abundant small mammals in their native range. Biol. Conserv. 144, 1335–1346. doi: 10.1016/j.biocon.2011.02.012
Derner, J. D., Detling, J. K., and Antolin, M. F. (2006). Are livestock weight-gains affected by black-tailed prairie dogs? Front. Ecol. Environ. 9:459–464. doi: 10.1890/1540-929520064[459:ALWGAB]2.0.CO;2
Detling, J. K. (2006). “Do prairie dogs compete with livestock?,” in Conservation of the Black-Tailed Prairie dog: Saving North America’s Western Grasslands, ed. J. L. Hoogland (Washington, DC: Island Press), 66–92.
Eldridge, D. J., Bowker, M. A., Maestre, F. T., Roger, E., Reynolds, J. F., and Whitford, W. G. (2011). Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol. Lett. 14, 709–722. doi: 10.1111/j.1461-0248.2011.01630.x
Estes, J. A., and Palmisano, J. F. (1974). Sea otters: their role in structuring nearshore communities. Science 185, 1058–1060. doi: 10.1126/science.185.4156.1058
Fahong, Y., Li, S., Kilpatrick, W. C., McGuire, P. M., He, K., and Wei, W. (2012). Biogeographical study of plateau pikas Ochotona curzoniae (Lagomorpha, Ochotonidae). Zool. Sci. 29, 518–526. doi: 10.2108/zsj.29.518
Forrest, S. C., and Luchsinger, J. C. (2006). “Past and current chemical control of prairie dogs,” in Conservation of The Black-Tailed Prairie dog: Saving North America’s Western Grasslands, ed. J. L. Hoogland (Washington, DC: Island Press), 115–128.
Fulbright, T., Davies, K. W., and Archer, S. R. (2018). Wildlife responses to brush management: a contemporary evaluation. Rangeland Ecol. Manag. 71, 35–44. doi: 10.1016/j.rama.2017.07.001
Glendening, G. E., and Paulsen, H. A. (1955). Reproduction and Establishment of Velvet Mesquite As Related to Invasion of Semi-Desert Grasslands. Technical Bulletin 1127. Washington, DC: U.S. Department of Agriculture.
González-Roglich, M., Swenson, J. J., Villarreal, D., Jobbágy, E. G., and Jackson, R. B. (2015). Woody plant-cover dynamics in Argentine savannas from the 1880s to 2000s: the interplay of encroachment and agriculture conversion at varying scales. Ecosystems 18, 481–492. doi: 10.1007/s10021-015-9841-5
Gordon, C. E., and Letnic, M. (2019). Evidence that the functional extinction of small mammals facilitates encroachment following wildfire in arid Australia. J. Arid Environ. 164, 60–68. doi: 10.1016/j.jaridenv.2019.01.015
Gori, D., and Schussman, H. (2005). State of the Las Cienegas National Conservation Area. Part I. Condition and Trend of the Desert Grassland and Watershed. Phoenix, AZ: The Nature Conservancy of Arizona, 63.
Gori, D. F., and Enquist, C. A. F. (2003). An Assessment of the Spatial Extent and Condition of Grasslands in Central and Southern Arizona, Southwestern New Mexico and Northern Mexico. Phoenix, AZ: The Nature Conservancy Arizona Chapter, 28.
Guyton, J. A., Pansu, J., Hutchinson, M. C., Kartzinel, T. R., Potter, A. B., Coverdale, T. C., et al. (2020). Trophic rewilding revives biotic resistance to shrub invasion. Nat. Ecol. Evol. 4, 712–724. doi: 10.1038/s41559-019-1068-y
Hale, S. L., and Koprowski, J. L. (2018). Ecosystem-level effects of keystone species reintroduction: a literature review. Restor. Ecol. 26, 439–445. doi: 10.1111/rec.12684
Hale, S. L., Koprowski, J. L., and Hicks, H. (2013). “Review of black-tailed prairie dog reintroduction strategies and site selection: Arizona reintroduction,” in Proceedings of the Merging Science and Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago III, 1-5 May 2012, Tucson, AZ, eds G. J. Gottfried, P. F. Folliott, B. S. Gebow, L. G. Eskew, and L. C. Collins (Fort Collins, CO: U.S. Department of Agriculture), 310–315.
Hamilton, W. T., McGinty, A., Ueckert, D. N., Hanselka, C. W., and Lee, M. R. (2004). Brush Management: Past, Present, Future. College Station, TX: Texas A&M University Press.
Hoogland, J. L. (1995). The Black-Tailed Prairie dog. Social Life of a Burrowing Mammal. Chicago, IL: University of Chicago Press.
Jackson, J. E., Branch, L. C., and Villarreal, D. (1996). Lagostomus maximus. Mamm. Species 543, 1–6.
King, J. A. (1955). Social Behavior, Social Organization, and Population Dynamics in a Black-Tailed Prairie dog Town in the Black Hills of South Dakota. Contributions from the Laboratory of Vertebrate Biology. Ann Arbor, MI: University of Michigan.
Kotliar, N. B., Baker, B. W., Whicker, A. D., and Plumb, G. (1999). A critical review of assumptions about the prairie dog as a keystone species. Environ. Manage. 24, 177–192. doi: 10.1007/s002679900225
Kotliar, N. B., Miller, B. J., Reading, R. P., and Clark, T. W. (2006). “The prairie dog as a keystone species,” in Conservation of the Black-Tailed Prairie dog: Saving North America’s Western Grasslands, ed. J. L. Hoogland (Washington, DC: Island Press), 53–64.
Krueger, K. (1986). Feeding relationships among bison, pronghorn, and prairie dogs: an experimental analysis. Ecology 67, 760–770. doi: 10.2307/1937699
Luce, R. J., Manes, R., and Van Pelt, B. (2006). “A multi-state plan to conserve prairie dogs,” in Conservation of the Black-Tailed Prairie dog: Saving North America’s Western Grasslands, ed. J. L. Hoogland (Washington, DC: Island Press), 210–217.
Miller, B., Ceballos, G., and Reading, R. (1994). The prairie dog and biotic diversity. Conserv. Biol. 8, 677–681. doi: 10.1046/j.1523-1739.1994.08030677.x
Miller, B., Reading, R., Hoogland, J., Clark, T., Ceballos, G., List, R., et al. (2000). The role of prairie dogs as keystone species: response to Stapp. Conserv. Biol. 14, 318–321. doi: 10.1046/j.1523-1739.2000.99201.x
Milne-Laux, S., and Sweitzer, R. A. (2006). Experimentally induced colony expansion by black-tailed prairie dogs (Cynomys ludovicianus) and implications for conservation. J. Mammal. 87, 296–303. doi: 10.1644/05-mamm-a-056r2.1
Noble, J. C., Hik, D. S., and Sinclair, A. R. E. (2007). Landscape ecology of the burrowing bettong: fire and marsupial biocontrol of shrubs in semi-arid Australia. Rangeland J. 29, 107–119. doi: 10.1071/RJ06041
O’Meilia, M. E., Knopf, F. L., and Lewis, J. C. (1982). Some consequences of competition between prairie dogs and beef cattle. J. Range Manage. 35, 580–585.
Ponce-Guevara, E., Davidson, A., Sierra-Corona, R., and Ceballos, G. (2016). Interactive effects of black-tailed prairie dogs and cattle on shrub encroachment in a desert grassland ecosystem. PLoS One 11:e0154748. doi: 10.1371/journal.pone.0154748
Proctor, J., Haskins, B., and Forrest, S. C. (2006). “Focal areas for conservation of prairie dogs and the grassland ecosystem,” in Conservation of the Black-Tailed Prairie dog: Saving North America’s Western Grasslands, ed. J. L. Hoogland (Washington, DC: Island Press), 232–247.
Reeve, A. F., and Vosburgh, T. C. (2006). “Recreational shooting of prairie dogs,” in Conservation of the Black-Tailed Prairie dog: Saving North America’s Western Grasslands, ed. J. L. Hoogland (Washington, DC: Island Press), 139–156.
Reisewitz, S. E., Estes, J. A., and Simenstad, C. A. (2006). Indirect food web interactions: seaotters and kelp forest fishes in the Aleutian archipelago. Oecologia 146, 623–631. doi: 10.1007/s00442-005-0230-1
Sankaran, M., Augustine, D. J., and Ratnam, J. (2013). Native ungulates of diverse body sizes collectively regulate long-term woody plant demography and structure of a semi-arid savanna. J. Ecol. 101, 1389–1399. doi: 10.1111/1365-2745.12147
Smith, A. T., and Foggin, J. M. (1999). The plateau pika (Ochotona curzoniae) is a keystone species for biodiversity on the Tibetan plateau. Anim. Conserv. 2, 235–240. doi: 10.1111/j.1469-1795.1999.tb00069.x
Staver, A. C., Bond, W. J., Stock, W. D., van Rensburg, S. J., and Waldram, M. S. (2009). Browsing and fire interact to suppress tree density in an African savanna. Ecol. Appl. 19, 1909–1919. doi: 10.1890/08-1907.1
Underwood, J. G., and Van Pelt, W. E. (2000). A Proposal to Reestablish the Black-Tailed Prairie dog (Cynomys ludovicianus) to Southern Arizona. Nongame and Endangered Wildlife Program Draft Technical Report. Phoenix, AZ: Arizona Game and Fish Department.
Vermeire, L. T., Heitschmidt, R. K., Johnson, P. S., and Sowell, B. F. (2004). The prairie dog story: do we have it right? Bioscience 54, 689–695.
Weltzin, J. F., Archer, S. R., and Heitschmidt, R. K. (1997). Small-mammal regulation of vegetation structure in a temperate savanna. Ecology 78, 751–763. doi: 10.1890/0012-9658(1997)078[0751:smrovs]2.0.co;2
Keywords: black-tailed prairie dog, browse trap, ecosystem engineer, exclosure, grassland, keystone species, reintroduction, woody plant encroachment
Citation: Hale SL, Koprowski JL and Archer SR (2020) Black-Tailed Prairie Dog (Cynomys ludovicianus) Reintroduction Can Limit Woody Plant Proliferation in Grasslands. Front. Ecol. Evol. 8:233. doi: 10.3389/fevo.2020.00233
Received: 30 January 2020; Accepted: 26 June 2020;
Published: 17 July 2020.
Edited by:
Peter Convey, British Antarctic Survey (BAS), United KingdomReviewed by:
John Lytton Hoogland, University of Maryland Center for Environmental Science (UMCES), United StatesCopyright © 2020 Hale, Koprowski and Archer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sarah L. Hale, c2hhbGUxNkBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.