 Boyang Sun1,2
Boyang Sun1,2 Tao Deng1,2,3*
Tao Deng1,2,3*- 1Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China
- 2CAS Center for Excellence in Life and Paleoenvironment, Beijing, China
- 3University of Chinese Academy of Sciences, Beijing, China
To approach a comprehensive understanding of the Equus Datum, we summarize the history of the study of Chinese fossil equids, from the “dragon bones” stage to scientific investigation, by Chinese, European and American people, and then review the stenonid species in China and discuss the phylogenetic relationships of early Equus. We conclude that there were at least two dispersal waves of Equus into the Old World from North America, and the second wave led to a radiation of the genus Equus in China. In China, the ages of the lower boundaries of the fossiliferous layers in classical Early Pleistocene faunal localities are all around 2.5 Ma (close to the age of the lower boundary of the Quaternary) according to magnetostratigraphic and biostratigraphic analyses. All of the geochronological results reveal China as an evolutionary center of Equus in Eurasia. In the late Neogene, the dominant equid lineage in China was Hipparionini. Even in the Pliocene, when hipparionines were not as widespread as before, there were still 5 genera and 7 species. In the Early Pleistocene, the hipparionines included only 2 genera and 2 species. In the meantime, global cooling caused Arctic ice sheets to form, lowering the sea level so that the Bering land bridge fell dry and allowed Equus to disperse into China. The rise of Equus was accompanied by the recession of hipparionines. The dispersal and radiation of Equus in China were not a coincidence, but a complicated process of adaptation and competition, and a comprehensive outcome of significant climatic, tectonic and biotic events.
Introduction
The Equus Datum, the first appearance of Equus in the Old World, was a remarkable biotic event in the Quaternary. It has often been used as a marker of the lower boundary of the Quaternary. This biotic event was contemporary with other important geologic and environmental events. Equus fossils are critical research material for studies on evolutionary, climatic, and environmental changes during the Quaternary in Eurasia (Deng and Xue, 1999a). Therefore, fossil Equus are essential for elucidating the Pleistocene paleobiology and paleoenvironment in Eurasia.
Chinese people were the first to obtain and utilize Equus fossils. Since historical times in China, vertebrate fossils, which were called “dragon bones,” have been used as pharmaceutical material. The “dragon bones” include Equus skeleton fossils. Fossil teeth of Equus are also regarded as pharmaceutical material and called “dragon teeth.” To review the history of paleontological research in Yushe Basin, Shanxi Province, Qiu and Tedford (2013) investigated historical records of “dragon bones” in the Shanxi area from ancient Chinese literature. They consulted Li Shizhen's famous “Compendium of Materia Medica” and found that the first reliable record of “dragon bones” as pharmaceutical material was in a lost medical book written by Lei Xue, who lived in the Song Dynasty (420–479 A.D.) of the Southern Dynasties. According to Tao Hongjing (456–536 A.D.), who lived in the Liang Dynasty of the Southern Dynasties, the Shanxi area was listed as the only place of origin for “dragon bones.” Finally, they concluded that the earliest reliable history of “dragon bones” can probably be traced to the fifth century, at least for Shanxi Province.
However, actual recognition of fossil Equus still depends on scientific observations and descriptions, which were initiated hundreds of years later by Western scholars. Matsumoto (1915) erected a new species, E. leptostylus, based on an isolated upper premolar from the Yinxu site, although this species was soon after considered invalid. However, the critical specimens for the Equus Datum date to the Early Pleistocene, which is when Equus invaded Eurasia. Later, they were called stenonid horses (see below). Vertebrate paleontologists from Europe and the United States began to study early Equus from China in the early twentieth century. Teilhard de Chardin and Piveteau (1930) reported the classic Pleistocene mammalian fauna, the Nihewan Fauna from Yangyuan, Hebei Province, and erected Equus sanmeniensis, which was the first named Chinese stenonid species. Zdansky (1935) first published a monograph of Equus in China based on the abundant specimens of the Lagrelius Collection housed in Uppsala, which were collected from Hebei, Henan, Shanxi, and Shandong by Andersson. Zdansky identified all stenonid specimens in this fossil series as Equus cf. sanmeniensis. Hopwood (1936) divided Eurasian Equus into caballines and zebrines. Early Equus species were considered zebrines, because they had a V-shaped linguaflexid like extant zebras. Colbert (1940) studied the Equus specimens collected from Yuanmou, Yunnan, by the Central Asiatic Expeditions research team of the American Museum of Natural History and erected Equus yunnanensis. He concluded that this new species was identical to the Pleistocene Equus from the upper Irrawaddy of Myanmar and was similar to the Pleistocene Equus of India in certain features. Eisenmann (1975) rediscovered E. sanmeniensis specimens in the Nihewan Fauna, which was divided into two different forms based on size by Teilhard de Chardin and Piveteau (1930). She erected a new species, Equus teilhardi, for the small form, and argued that in addition to its small size, its lack of an incisor cup distinguished it from other Equus species.
After 1949, as the economy was rebuilt and science progressed, Chinese scholars began to research early Equus. Chow and Liu (1959) described cheek tooth specimens of Equus collected from Pinglu, Shanxi Province, and erected a new species, Equus huanghoensis. Pei (1961) reported new specimens of E. yunnanensis collected from Yuanmou, Yunnan Province, and Gigantopithecus Cave in Liucheng, Guangxi Province. Liu (1973) reported a mandible specimen of Equus in Locality 1 of Zhoukoudian; he identified it as E. sanmeniensis and thus extended the last occurrence of this species to the Middle Pleistocene. Liu and You (1974) described much better specimens of E. yunnanensis than previously discovered, which included a skull and mandible. Forsten (1984) proposed a “stenonid pattern” to describe the V-shaped linguaflexid when she discussed the supraspecific classification of the Old World hipparionines. The term “stenonid” references E. stenonis, which was the first erected species among early Equus, and gradually came to be used as a generic term for early Equus, especially by the authors of certain extensive review publications (Alberdi et al., 1998; Deng and Xue, 1999a).
Deng and Xue (1999b,c), respectively, erected two new species, Equus qingyangensis and Equus wangi, based on the specimens collected from Bajiazui, Qingyang, Gansu. In the same year, they also published the most comprehensive review of early Equus, or stenonids: the monograph Chinese Fossil Horse of Equus and Their Environment (Deng and Xue, 1999a). Qiu et al. (2004) reported a new Pleistocene mammalian fauna, the Longdan Fauna, from Dongxiang, Gansu Province. They erected Equus eisenmannae based on the large equid form in this fauna. Dong and Fang (2005) reported some fragmented cranial specimens of E. huanghoensis found from Tangshan, Nanjing, Jiangsu Province, and supplied some important features of E. huanghoensis. Li et al. (2016) reported the most complete fossil record yet known of E. huanghoensis, which included a skull associated with a mandible from Yangshuizhan in Yangyuan, Hebei Province. Sun et al. (2017) rediscovered a skull and mandible in the Lagrelius Collection from Qixian, Shanxi Province, which was previously identified as E. cf. sanmeniensis by Zdansky (1935). They identified the skull and mandible, respectively, as E. teilhardi and E. qingyangensis.
In this article, we will review all the known stenonid species on the robust basis of our predecessors. We will reorganize all important information about these species, explore their early evolution, and further elucidate the Equus Datum.
Materials and Methods
For phylogenetic analysis, we reorganized the data matrices from previous publications and summarized them in a new data matrix, which includes 16 taxa, and 53 characters. The phylogenetic analysis were performed using TNT 1.1 with a traditional search method (Goloboff et al., 2008), 1,000 replications and the trees-bisection-reconnection branch-swapping algorithm (TBR) applied in our analysis. All characters are equally weighted and additive. Gaps are treated as “missing” and multistate taxa interpreted as polymorphism. The resulting strict consensus tree is presented in Figure 1. Character list and data matrix are presented in the Supplementary Information.
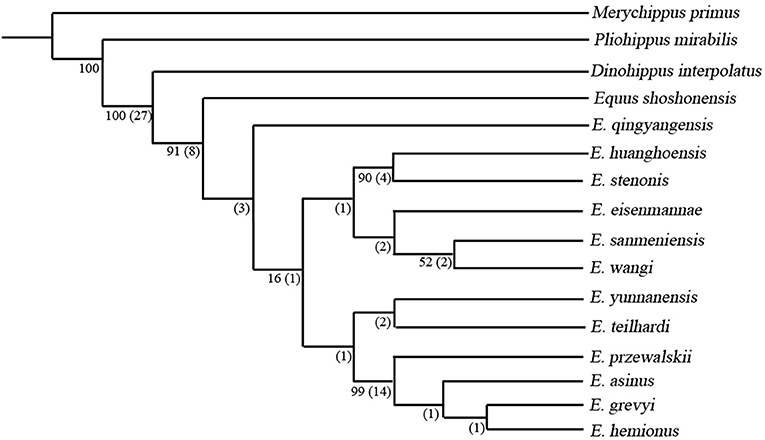
Figure 1. Most parsimonious tree (MPT) obtained from phylogenetic analysis, Tree length = 148, consistency index = 0.635, retention index = 0.768. Numbers by the nodes denote the Bootstrap (outside brackets, not shown if value <50) and Bremer values (in brackets, not shown if value <1).
Systematic Paleontology of Stenonids in China
We review all of the stenonids species in China in the order in which they were erected. All discussions are based on the classical descriptions and the most recent reports. The main localities of stenonids are shown in Figure 2.
Order Perissodactyla Owen, 1848
Family Equidae Gray, 1821
Genus Equus Linnaeus, 1758
Equus stenonis Cocchi, 1867
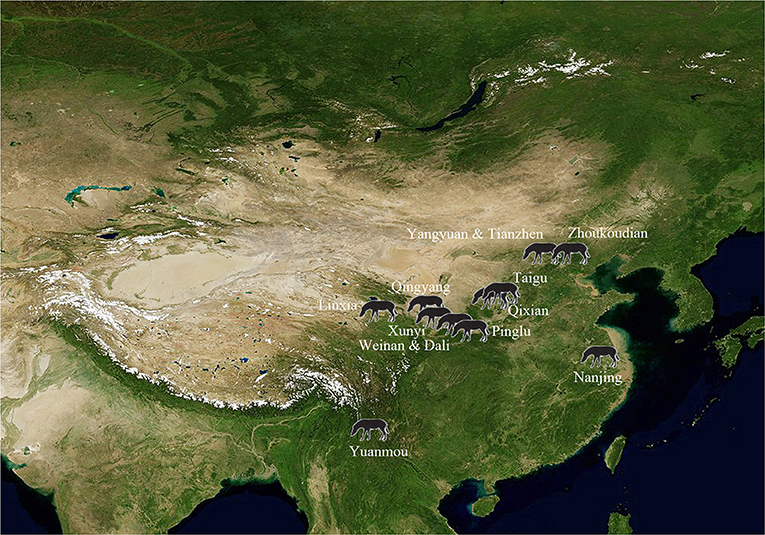
Figure 2. Main localities of stenonids in China.
Holotype: IGF−560, adult skull with mandible, collected from the Lower Pleistocene of the upper Arno valley in Italy.
Diagnosis (following Deng and Xue, 1999a): large size, facial angle is very narrow, well-developed preorbital pit retained, deep groove along nasal suture, lateral outline of skull undulated, domed parietal, frontal also undulated laterally. DP1 present, diastema elongated, protocone short, hypocone extends lingually, plications complex. Linguaflexid sharply V-shaped, cup on lower incisor developed. Snout elongated, nasal notch very deep, its posterior margin is at the level of mesostyle or even the posterior margin of P3, and the anterior foramen of the palatine canal is open to the level of M3.
Age: Nihewanian, Early Pleistocene.
Distribution: Eurasia and North America.
Remarks: Equus stenonis was the first erected stenonid species (Figure 3). This species has been studied for a long time, and there is an abundance of specimens. Its fossil record is widespread in Eurasia, and there are many recognized subspecies, although some of them have been upgraded to species, such as E. livenzovensis (Alberdi et al., 1998). However, a definite record of E. stenonis in China was obtained late.

Figure 3. Right lateral view of cast of holotype skull with mandible of E. stenonis, NHMUK PV M 29511.
Azzaroli (1982) observed a skull collected from Taigu, Shanxi Province, that is housed in the Frick Collection in the AMNH. He identified this skull as E. stenonis. This is the earliest formal report of E. stenonis in China. Forsten (1986) studied the specimens collected from Taigu that are housed in the Licent Collection in the THP. She argued that the protocones of the Taigu specimens were longer than E. stenonis material from any locality in Europe, so the Frick specimen should not be considered this species. Azzaroli (1987) argued that the locality information of the Licent specimens was not clear and not comparable to the Frick specimen. Deng and Xue (1999a) observed the Frick specimen based on plate published by Azzaroli (1982) and recognized that many features indeed belonged to E. stenonis.
Azzaroli et al. (1988) proposed that E. stenonis dispersed into Europe about 2 Ma and survived until the end of the Early Pleistocene at 1 Ma. Deng and Xue (1999a) considered E. stenonis to be the probable ancestor of E. stehlini and E. bressanus, which are both common fossil equids found in Europe. Deng and Xue (1999a) described a molar of E. stenonis from Xinyaozi Fauna in Tianzhen, Shanxi and they noted that the age of the Xinyaozi Fauna was older than that of the Nihewan Fauna s. s. Azzaroli and Voorhies (1993) proposed that the North American subspecies E. stenonis anguinus occurred about 2 Ma. Therefore, the earliest record of E. stenonis known so far is in China, which was probably the center of origin of this species.
Equus sanmeniensis Teilhard de Chardin and Piveteau, 1930
Lectotype: NIH 002 (Figure 4), aged adult skull with mandible, collected from Nihewan, Yangyuan, Hebei Province, formally assigned by Eisenmann (1975).

Figure 4. Right lateral view of lectotype skull of E. sanmeniensis, NIH 002, collected from Nihewan Basin.
Diagnosis (following Deng and Xue, 1999a): giant build, skull elongated, basal narrow and short so it shown an elongated snout with narrow frontal, DP1 stably present. Protocone elongated and concave lingually, labial margins of para- and mesostyle often concave, pli caballine strong, but often diminished to absent by wear, M3 has isolated enamel ring and double-angled occlusal surface. Linguaflexid V-shaped, ectoflexid deep, penetrating into the isthmus, even touching the linguaflexid on lower molars, making lingual margin of linguaflexid and labial margin of ectoflexid flat. Plications complex, but often diminished to absent by wear.
Age: Nihewanian–Zhoukoudianian, Early to Middle Pleistocene.
Distribution: North and Northwest China, Siberia, probably Tajikistan.
Remarks: Teilhard de Chardin and Piveteau (1930) erected the first stenonid species in China, E. sanmeniensis. This species was distributed in northern China, Siberia, and Tajikistan from the Early to Middle Pleistocene (Deng and Xue, 1999a). This species evolved early, and was an outcome of the diversification of Equus that dispersed into Eurasia approximately 2.5 Ma (see below). Equus sanmeniensis has primitive features similar to the earliest Equus species in North America, E. shoshonensis, such as a transversely undulated lateral outline of skull, groove along the nasal suture, elongated facial part, and obvious preorbital pits. However, E. sanmeniensis differs from E. stenonis by having a less deep nasal notch and an elongated protocone. All of these features indicate that E. sanmeniensis and E. stenonis share derived features inherited from some common ancestor type, which most likely lived in North America, but they evolved along different trajectories. Therefore, we conclude that E. sanmeniensis and E. stenonis underwent parallel evolution, and they have a close but not an ancestor-descendant relationship. Early on, authors tended to identify more stenonid specimens as E. sanmeniensis. Teilhard de Chardin and Piveteau (1930) described all Equus species they found in the Nihewan Fauna as E. sanmeniensis, even though they noticed that there were two different types of specimens. Zdansky (1935) identified all of the stenonid specimens in his study as E. cf. sanmeniensis. Some of these have been revised (Eisenmann, 1975; Sun et al., 2017); however, we believe that lots of specimens of other stenonid species are still considered E. sanmeniensis.
Equus yunnanensis Colbert, 1940
Lectotype (assigned by Deng and Xue, 1999a): IVPP V 4250.1, female skull, about 7–8 years old, extrusion-deformed, well-preserved except for a little damage.
Paralectotype: IVPP V 4251, senile male mandible, coronoid process damaged. All of type specimens are collected from Madahaixigou, Youmou, Yunnan.
Diagnosis (following Deng and Xue, 1999a): Medium size, skull relatively large, frontal narrow, snout elongated. Lingual margin of I3 not enclosed, size of cheek teeth and length of protocone medium, enamel plications usually strong, linguaflexid V-shaped, metaconid and metastylid very rounded, ectoflexid shallow, rarely penetrates into the isthmus even on lower molars, pli caballinid not strong, posterior tip of m3 halberd-shaped. Distal limb robust.
Age: Nihewanian to Salawusuan, Early to Late Pleistocene.
Distribution: Yuanmou, Yunnan; Liucheng, Guangxi; Jianshi and Enshi, Hubei; Hanzhong, Shaanxi; Huili, Sichuan; Irawaddy, Myanmar.
Remarks: Colbert (1940) erected E. yunnanensis based only on isolated cheek teeth. Therefore, it was difficult for researchers to conduct enough work on morphology of this species. Fortunately, Liu and You (1974) discovered and reported well-preserved cranial and limb specimens from Yuanmou Basin. They observed much more morphological information than Colbert (1940) and further discussed the relationships of Asian Equus. They argued that E. yunnanensis should be considered a separate lineage distinct from other species in South Asia. They suggested that Equus sp. found in Huili, Sichuan Province, was likely derived from E. yunnanensis. Therefore, this lineage seemed to have dispersed to the north. Deng and Xue (1999a) thought that E. yunnanensis and E. wangi were closely related because they shared similar derived features, such as rounded double-knots and short ectoflexids. Pei (1961) noted that E. yunnanensis was an equid species similar to asses and horses based on its short ectoflexid. Deng and Xue (1999a) argued that short ectoflexids should not be thought to be particular to asses, because other equids, such as E. wangi and E. przewalskii, also have this feature. Based mainly on rounded double-knots and short ectoflexids, Deng and Xue (1999a) considered these species sister-groups. Gromova (1949) and Deng and Xue (1999a) both suggested that the different dental features, such as the morphology of enamel plications on the upper cheek teeth, were adaptations to different environments. As an increasing number of specimens have been discovered, the arguments regarding phylogenetic relationships were revised in recent years. We propose some new ideas about the relationship between E. yunnanensis and other stenonid species below.
Equus huanghoensis Chow and Liu, 1959
Holotype: IVPP V 2385–2389, three upper premolars and two upper molars, collected from bank of Yellow River (Huanghe) in Sanmenxia, Pinglu, Shanxi Province.
Diagnosis (following Li et al., 2016): Large skull with elongated face and snout; outline of parietal part undulated as seen in lateral view. Nasal-frontal-parietal part with hollow in middle part and upturned anterior end, middle groove of nasal bone narrow. Nasal notch deepens to reach level of boundary of P3/P4. Orbit posterior of cheek teeth row, anterior foramen of palatine canal opens posteriorly; interalveolar margin long. Occipital plane pentagonal, and supramagnum protuber strong. Protocone shortened, mesostyle robust, pli caballine very weak, and plication simple. Mandible elongated. Condyle of mandible elongated transversely with rounded anterior margin and cupped internally posterior margin. Cups of lower i3 enclosed, pli caballinid weak to absent, enamel plication of post-flexid strong, and linguaflexid V-shaped. Locality and age: Nihewanian, Early Pleistocene. Distribution: Nihewan, Hebei; Linyi, Shanxi; Sanmenxia, Pinglu, Shanxi; Xunyi, Shaanxi; Nanjing, Jiangsu.
Remarks: Chow and Liu (1959) erected Equus huanghoensis based on five isolated upper cheek teeth collected from Sanmenxia, Pinglu, Shanxi Province. Forsten (1986) reviewed numerous classical specimens of Chinese fossil Equus. She identified a skull from the Lagrelius Collection in the Museum of Evolution, Uppsala University, as? Equus huanghoensis based on the very short protocone of this specimen. Deng and Xue (1999a) described E. huanghoensis specimens from Xunyi, Shaanxi Province, and stated that E. huanghoensis is primitive because of its very short protocone, and further identified this species as a stenonid. Dong and Fang (2005) reported some fragmented cranial material of E. huanghoensis from Tangshan, Nanjing, Jiangsu Province. Li et al. (2016) reported the most complete material ever known of E. huanghoensis from Nihewan, Yangyuan, Hebei Province, which included a complete skull with a broken mandible, and described all of the critical features of this species (Figure 5). Equus huanghoensis is a very large stenonid species with deep nasal notch, lower cranial proportion, elongated snout and facial part, and V-shaped linguaflexid. All of these features supported Deng and Xue's argument that E. huanghoensis is a stenonid species. Based on the measurements of Nihewan material, E. huanghoensis is one of the Equus species with the largest absolute and relative palatal lengths, and is similar to E. eisenmannae, which was the largest Equus species in China (Qiu et al., 2004; Figure 6). Deng and Xue (1999a) suggested that the age of E. huanghoensis was in the Early Nihewanian (2.5–1.8 Ma). Ao et al. (2013b) suggested that the age of the E. huanghoensis specimens from Nihewan was around 1.7 Ma. This is the youngest record of E. huanghoensis ever known in China. In addition, the Nihewan material is smaller compared with the holotype from Sanmenxia. Li et al. (2016) indicated that the P4 and M2 lengths of the holotype are around 1.16 times larger than those of the Nihewan specimen. The ratio between the length of the basilar and cheek tooth row is generally stable in Equus (Figure 7). Based on this ratio, the holotype teeth should belong to a very large individual whose basilar length was 647.28 mm, in contrast with that of E. eisenmannae (625 mm, Qiu et al., 2004).
Equus teilhardi Eisenmann, 1975

Figure 5. Left lateral view of skull of E. huanghoensis, NWUV 1403.1, collected from Yangshuizhan Locality, Nihewan Basin. Scale bar = 5 cm.
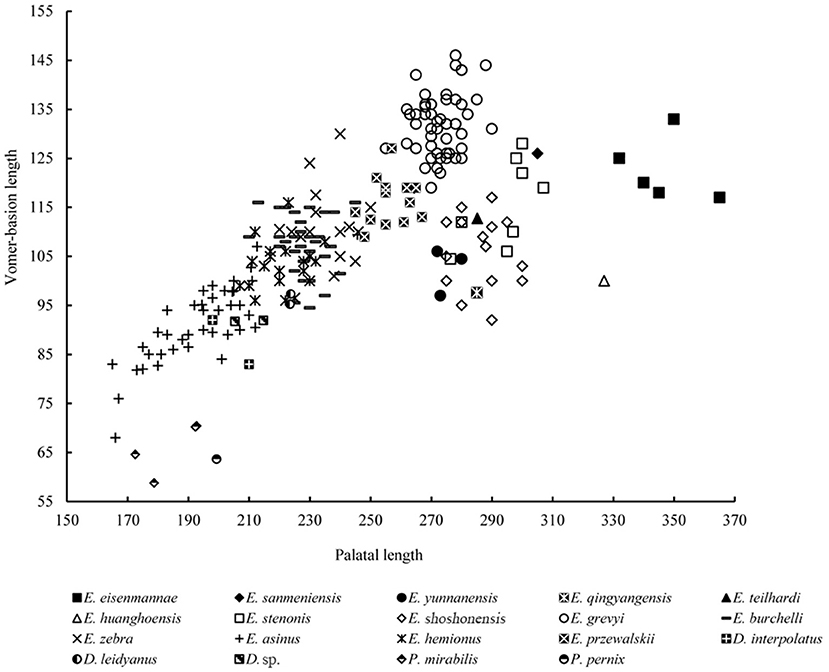
Figure 6. Comparison of lengths of palatine and basicranial in several extinct and extant species of Equidae. Abbreviations of genera: E, Equus; D, Dinohippus; P, Pliohippus (same in below).
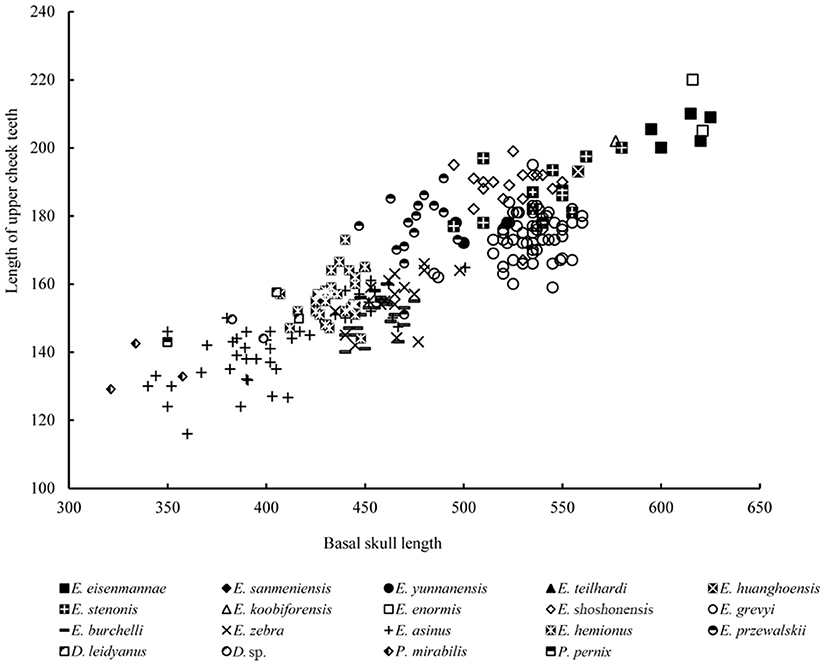
Figure 7. Comparison of cheek tooth row length and basilar length in several extinct and extant species of Equidae.
Holotype: NIH 001, a broken mandible with symphysis and left ramus.
Diagnosis (following Sun et al., 2017): Middle size. The nasal notch extends above the mid-P2 to the boundary of P2/P3. Snout short, preorbital fossa shallow with indistinct border, lateral outline is undulated, deep groove along nasal suture. Cups of incisors possibly incomplete or even lacking. The upper cheek teeth have simple fossettes, short protocones and weak plis caballines. The lower cheek teeth have V-shaped linguaflexids and deep ectoflexids on molar that penetrate into the isthmuses, even touching the linguaflexids on the lower molars. The limb bones are short and robust. Distribution: Northwestern and northern China. Age: Nihewanian, Early Pleistocene.
Remarks: When Teilhard de Chardin and Piveteau (1930) erected E. sanmeniensis, they recognized two forms of Nihewan specimens with different body sizes. However, they did not resolve this issue and still identified all specimens as E. sanmeniensis. In Zdansky's study on Chinese fossil Equus, he described many specimens with different features and from different localities as Equus cf. sanmeniensis. Later, when Eisenmann studied Nihewan equids housed in Paris, she erected E. teilhardi based on a broken mandible. Eisenmann (1975) only listed two features in the diagnosis: small size and lack of lower incisor cup. Forsten (1986) studied the equid materials of the Nihewan fauna collected by Licent and identified two fragmented cranial specimens (THP 00150, 00151) as E. teilhardi. She proposed many important features for cranial and dental morphology. Deng and Xue (1999a) described a left ramus fragment (NWUV 1243) from Bajiazui, Qingyang, Gansu Province. They focused on the absence of a cup on the lower incisor of E. teilhardi and argued that this species should be regarded as a lineage distinct from other Eurasian stenonids. Sun et al. (2017) reviewed the equid specimens in the Lagrelius Collection described by Zdansky (1935) and recognized a skull from Qixian, Shanxi Province, as E. teilhardi (Figure 8). The features of this skull are basically identical to those described by Forsten. Based on measurements of Qixian material, Sun et al. (2017) noted that E. teilhardi had relatively shorter distal limbs than most Eurasian stenonids. Additionally, based on the I3 character of Qixian material and observation of other early Equus specimens, such as E. shoshonensis in the Hagerman Collection, Sun et al. (2017) argued that the lack of a lower incisor cup of E. teilhardi was due to variation. Some European stenonid horses, such as E. stehlini, also have incomplete lower incisor cups (Azzaroli, 1965, 1982). Based on skull and post-cranial morphology, Sun et al. (2017) hypothesized that E. teilhardi and E. yunnanensis were probably closely related.
Equus qingyangensis Deng and Xue, 1999b

Figure 8. Left lateral view of skull of E. teilhardi, PMU M 1321, collected from Qixian, Shanxi, housed in Museum of Evolution, Uppsala University. Scale bar = 5 cm.
Holotype: NWUV 1128, the middle and back of a skull with all cheek teeth, about 12 years. Diagnosis: Medium size. Nasal notch extends above back of P2, preorbital pit is shallow with indistinct border, deep groove along the nasal suture. The upper teeth have long protocones and weak plis caballines. The lower teeth have typical V-shaped linguaflexids and deep ectoflexids that penetrate into the isthmuses or even touch the linguaflexids on the lower molars. The limb bones are slender. The metacarpal middle shaft index is smaller than 13.5 and the metatarsal smaller than 12.0. It was the early species of Equus with the most slender limb bones in Eurasia. Distribution: Northwestern and northern China. Age: Nihewanian, Early Pleistocene.
Remarks: Deng and Xue (1999b) erected a new stenonid species, E. qingyangensis. According to their description, although E. qingyangensis is morphologically similar to E. sanmeniensis, except for the large body size of the latter, E. qingyangensis has some impressive features that have never been found in known stenonid horses in China. For example, its skull has about a 12 mm distance between the temporal condyle and the posterior margin of the orbital bar (Deng and Xue, 1999a). This is a primitive feature because of its presence in the hipparionine genus Proboscidipparion (Qiu et al., 1987), and in some primitive equid species, based on our observation, such as Pliohippus. In contrast, these two constructs are adjacent in most extinct and extant Equus species. The Mc III of E. qingyangensis is more slender than those of all known stenonid horses, with a middle shaft index of 13.3 (Deng and Xue, 1999a). Eisenmann and Deng (2005) collected measurements of E. qingyangensis, other Eurasian stenonids, and North American early Equus, and conducted a series of statistical analyses. They concluded that E. qingyangensis are similar to E. shoshonensis but different from E. stenonis based on cranial and post-cranial proportions. Our plot also shows that E. qingyangensis has very low ratio of cranial part (Figure 9), which has been regarded as a primitive feature (Qiu et al., 2004). Sun et al. (2017) found a similar phenomenon. Based on observation of the Hagerman Collection, they indicated that E. shoshonensis also have about a 10–12 mm distance between the temporal condyle and the posterior margin of the orbital bar, which is identical to that of E. qingyangensis. Eisenmann and Deng (2005) argued that E. shoshonensis and E. qingyangensis probably have a direct ancestor-descendant relationship and represent a lineage that is distinct from the Eurasian species E. stenonis. Sun et al. (2017) also hypothesized that E. qingyangensis may be a direct descendant of E. shoshonensis and the earliest and most primitive stenonid species in Eurasia.
Equus wangi Deng and Xue, 1999c
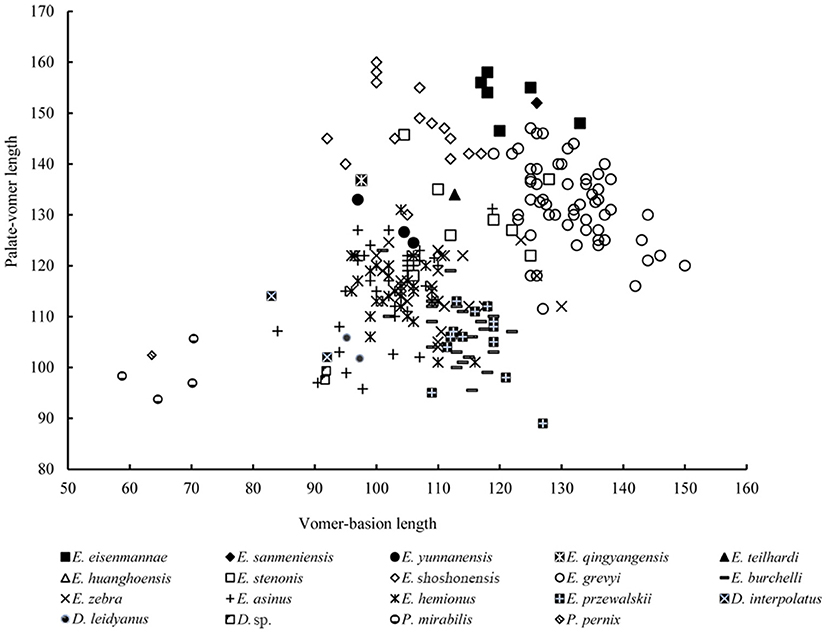
Figure 9. Comparison of basicranial proportion in several extinct and extant species of Equidea.
Holotype: NWUV 1170, complete upper and lower cheek tooth rows that belong to an individual, and the upper tooth rows go with the remains of the maxillae, about 5 years.
Diagnosis: Teeth are very large. DP1 present. Protocones long, narrow and middle-grooved. Mesostyles flat or lightly middle-grooved. Enamel plications strong, especially on the premolars. Hypoconal grooves on premolars with flat bottoms. The m3 has a double-angled back margin and an enamel lake within its hypocone. The double–knots are very circular so that their metastylids do not have posterior angles. Linguaflexids wide and sharply V-shaped. Ectoflexids shallow so that they never penetrate into isthmuses even on molars, and post-flexids correspondingly long. Plis caballinid much developed. Enamel plications of post-flexids strong. Entoconids on premolars with sharp beak-shaped ends. Hypoconulid on m3 like a short dagger with a wide and round end.
Distribution: Qingyang, Gansu.
Age: Nihewanian, Early Pleistocene.
Remarks: Deng and Xue (1999c) erected a new species, E. wangi, based on cheek tooth rows and a cranial fragment from Bajiazui, Qingyang, Gansu Province. They estimated that this species has a deep mid-nasal groove and preorbital pit based on the broken skull. The cheek tooth size of E. wangi is very large, similar to E. sanmeniensis, E. huanghoensis, and E. eisenmannae. However, E. wangi has obvious morphological differences from E. sanmeniensis and E. huanghoensis. Equus wangi has features in common with E. yunnanensis, such as circular metastylids and shallow ectoflexids (Deng and Xue, 1999a). Qiu et al. (2004) noted that E. eisenmannae has many characters in common with E. wangi, including large cheek teeth, short plis caballine, simple mesostyles, strong enamel plications, rounded double-knots, shallow ectoflexids, and elongated post-flexids. Another unusual feature of E. wangi is its beak-shaped entoconid. According to Qiu et al. (2004) and Wang and Deng (2011), E. eisenmannae also has similar lower premolar structures. Therefore, E. wangi is probably more closely related to E. eisenmannae than other stenonid species. In addition, E. wangi is similar to E. shoshonensis from Hagerman, Idaho, USA based on its very circular double-knot, beak-shaped premolar, and sharp V-shaped linguaflexid (Deng and Xue, 1999a). Equus shoshonensis was the earliest Equus species, whereas E. yunnanensis and E. eisenmannae occurred during the beginning of the Quaternary in China. Consequently, E. wangi should be a very primitive lineage. Deng and Xue suggested that first occurrence of E. wangi was between 2.5 and 2.4 Ma.
Equus eisenmannae Qiu et al., 2004
Holotype: IVPP V13552, a complete adult and male skull (Figure 10) with its mandible, about 12 years old.

Figure 10. Right lateral view of holotype skull of E. eisenmannae, IVPP V 13552, collected from Longdan Locality, Linxia Basin. Scale bar = 5 cm.
Diagnosis: (1) giant size with a basal skull length of 613.8 mm on average; (2) longer face than in any known horses; (3) shallow preorbital fossa; (4) high vomer index (111.2–133.9); (5) long distance between orbit and third molar; (6) rounded double-knots of lower cheek teeth; (7) shallow labial valleys on lower molars. Character states 2–6 are primitive within horses; character states 1 and 7 are considered derived.
Distribution: Longdan, Dongxiang County, Gansu Province.
Age: Nihewanian, Early Pleistocene.
Remarks: Equus eisenmannae was the most recently erected stenonid species in China. This species is very interesting. Based on measurement and comparison, Qiu et al. (2004) noted that E. eisenmannae was one of the largest Equus species (Figure 7). Furthermore, facial and snout lengths, and absolute and relative palatal lengths, were all larger than those of any other known Equus species (Figures 6, 11, 12). However, based on the method proposed by Eisenmann and Karchoud (1982), Qiu et al. (2004), and Sun et al. (2017) compared the Mc III and cranial basilar lengths among all Chinese stenonid horses and determined that E. eisenmannae has the largest relative robustness of Mc III. Equus eisenmannae also has the most abundant cranial material record among Chinese stenonids, including several complete skulls from different fossiliferous levels. Qiu et al. (2004) noticed that individuals from different levels have morphologic differences; for example, individuals from the upper level have larger cranial proportions, deeper ectoflexids in the lower molars, and sharper post-erolingual angles of the metastylid. Wang and Deng (2011) performed univariate and multivariate analyses on all specimens from different levels in the locality where E. eisenmannae was recovered. They concluded that the differences between E. eisenmannae samples across the two stratigraphic intervals were not significant enough to warrant recognizing two species. They also discussed some critical trends in equid evolution based on their comparison between E. eisenmannae and modern horses (Forsten, 1988; Azzaroli, 1992; Eisenmann, 1996; Prado and Alberdi, 1996; Eisenmann and Baylac, 2000). Qiu et al. (2004) summarized the important features of E. eisenmannae and indicated that its beak-shaped entoconids on the lower premolars and rounded double-knots were similar to those of E. wangi, and its oval entoconids on the lower molars and shallow labial valley not reaching the isthmus were similar to those of E. yunnanensis and E. wangi. They also argued that E. eisenmannae is a transitional form between North American plesippines and Eurasian allohippines (i.e., stenonids).
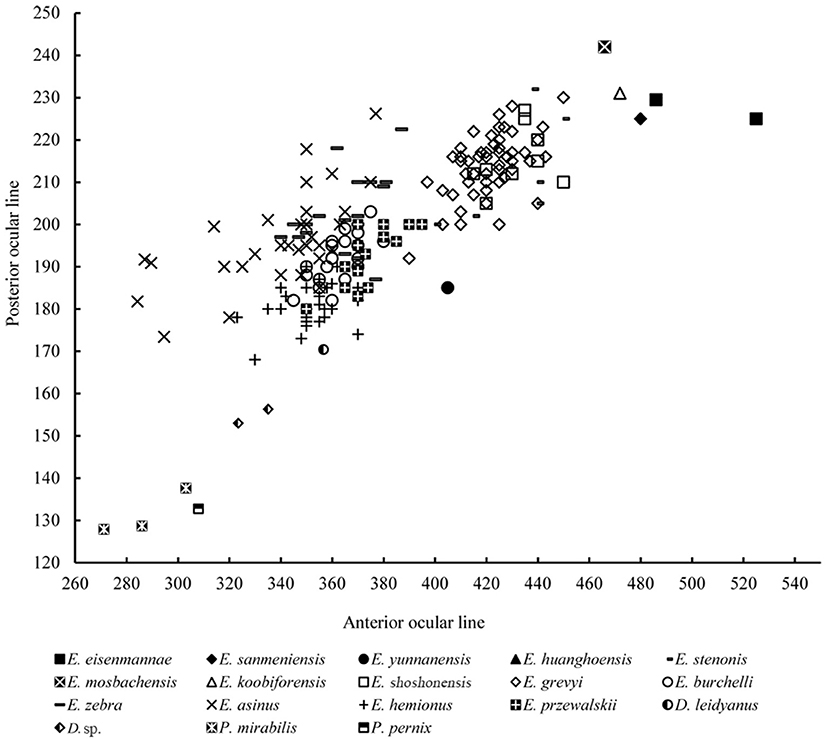
Figure 11. Comparison of anterior and posterior ocular lengths in several extinct and extant species of Equidae.
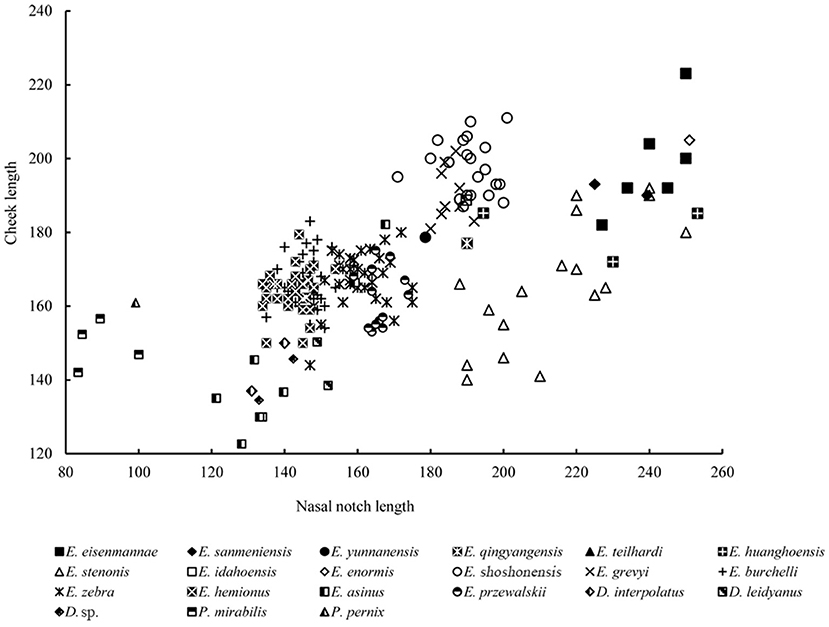
Figure 12. Comparison of lengths of nasal notch and cheek in several extinct and extant species of Equidae.
Summary of Chinese Stenonid Phylogenetic Relationships
Deng and Xue (1999d) reviewed all of the fossil equids in China and summarized their relationships in a phylogenetic tree. They proposed that E. teilhardi and E. huanghoensis are two independent lineages alone and traced these two species directly back to E. shoshonensis. They could not recognize diagnostic properties of these two species because of lack of complete material. Deng and Xue (1999a) considered the other stenonid species they reviewed to be a clade, within which E. qingyangensis was a separate lineage that they suggested directly dispersed into Eurasia as a New World origin. Equus wangi and E. yunnanensis are sister groups, as were E. stenonis and E. sanmeniensis. The establishment of these groups was also based on the morphological similarities we discussed above. As new lineages are discovered and more complete material of known species reported, it is necessary to perform a new review of the stenonid horses in China.
Some authors have suggested that Equus originated within genus Dinohippus, a large monodactyl form constrained to the New World (Bennett, 1980; Dalquest, 1988; MacFadden, 2005, 2006; Pagnac, 2006). In the current research, we try to figure out the origin of Equus in different method and then obtain different conclusions. In our plots, stenonids are more similar to Pliohippus in cranial proportion rather than to Dinohippus, caballoids, hemiones, asinines, and zebrines (Figures 6, 9, 11, 12). On the other hand, we perform a phylogenetic analysis. The analysis yield one parsimonious tree, shows a new phylogenetic relation between primitive monodactyl equids, stenonids, and some extant species (Figures 1, 13). In this phylogenetic tree, a monophyletic clade consists of species of Equus forms sister groups with species of Dinohippus. Moreover, the clade consists of Equus and Dinohippus forms sister groups with species of Pliohippus.
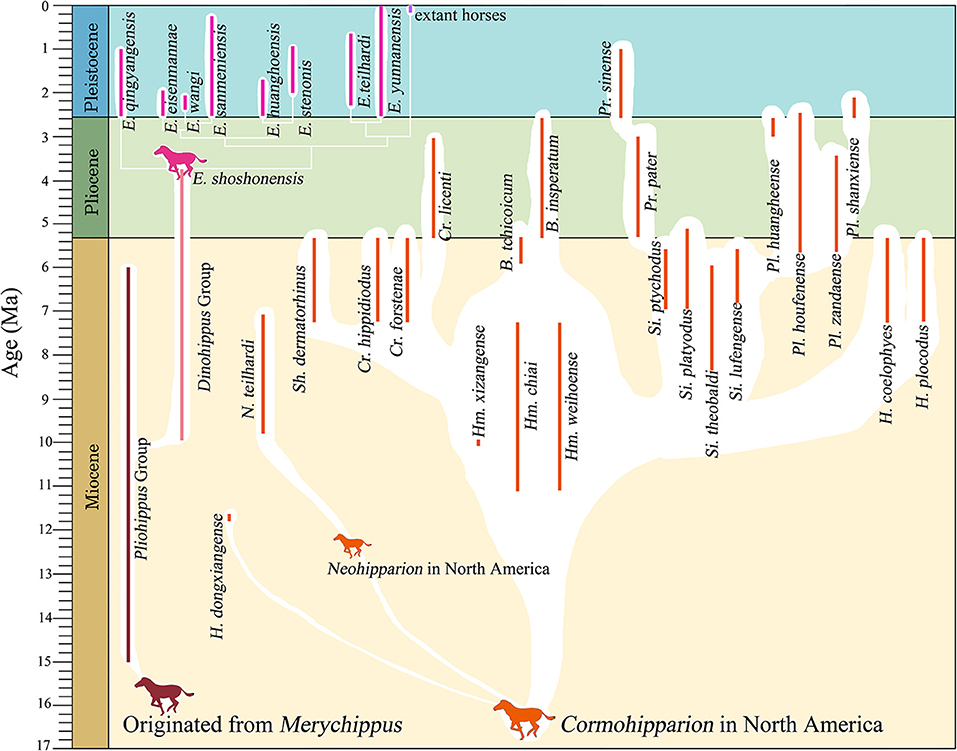
Figure 13. Proposed phylogenetic interrelationships of species of hipparionine and genus Equus in China, based on Qiu et al. (1987), Li and Li (1990), Deng and Xue (1999a), MacFadden (2005), Sun (2013), Wolf et al. (2013), Deng et al. (2016), Bernor et al. (2018), and Sun et al. (2018). Abbreviations of genera: H, Hipparion (a polyphyletic group including all the species with undefined attribution); N, Neohipparion; Sh, Shanxihippus; Cr, Cremohipparion; Hm, Hippotherium; B, Baryhipparion; Pr, Proboscidipparion; Si, Sivalhippus; Pl, Pliohipparion; E, Equus.
Eisenmann and Deng (2005) agreed with Deng and Xue's argument regarding the phylogenetic position of E. qingyangensis. Consequently, there were likely at least two dispersal waves of Equus into the Old World, with one wave represented by E. qingyangensis. Eisenmann and Deng (2005) argued that E. shoshonensis may be the direct ancestor of E. qingyangensis. In our result of new phylogenetic analysis, E. qingyangensis is attributed in a clade distinguished from all other species. This probably represents an independent dispersal event, and accords with the arguments of Deng and Xue (1999a) and Eisenmann and Deng (2005).
In the new phylogenetic tree, large-sized stenonid species form a clade. Large body size is a primitive character. Body size decrease was an important trend in equid evolution (Eisenmann and Baylac, 2000; Wang and Deng, 2011). The earliest Equus species in Europe, E. livenzovensis, is also very large, and is thought to have lived 2.5 Ma in Italy, Spain, and Russia. According to geochronological data, the first occurrences of E. huanghoensis and E. eisenmannae were close to the lower boundary of the Quaternary (see below). Although these huge species of Chinese stenonid have primitive body size and early geohistorical distribution, their features are derived. This probably suggested that the differentiation of early Equus was significantly earlier than 2.58 Ma. Qiu et al. (2004) considered E. eisenmannae to be the ancestor of E. sanmeniensis and E. stenonis. Based on morphology of cranial and dental features, E. eisenmannae is more closely related to E. sanmeniensis than E. stenonis. They share some morphological similarities, such as extremely large body size (basilar length of type skull of E. sanmeniensis is 585 mm, this length of E. eisenmannae is 613 mm), shallow nasal notch, elongated protocone, and strong enamel plications on upper cheek teeth. In addition, based on morphological features, E. wangi, E. sanmeniensis and E. eisenmannae probably have the closest relationship, fits the result of our phylogenetic analysis. Equus wangi and E. sanmeniensis formed a lineage as sister groups, which form a group with E. eisenmannae. Equus huanghoensis and E. stenonis shared features such as a transversely undulated forehead, very deep nasal notch, short protocone, and very weak enamel plications on upper cheek teeth. These similarities also fit the result of our phylogenetic analysis, in which Equus huanghoensis and E. stenonis form sister groups.
Previously, due to poor material, it was difficult to recognize the phylogenetic position of E. teilhardi (Eisenmann, 1975; Forsten, 1986). Recently, a report on a nearly complete skull of E. teilhardi provided a perfect opportunity to discuss this species (Sun et al., 2017). Most stenonid species have large body size, elongated facial part and limbs. In contrast, E. teilhardi has medium body size, a short snout and robust metapodials (Sun et al., 2017). E. yunnanensis has similar cranial and post-cranial features as E. teilhardi, such as body size, cranial proportions and limb shape. In the result of our phylogenetic analysis, E. teilhardi forms sister groups with E. yunnanensis. This result fits the similarities above in morphology feature. Due to the introduction of extant species (caballoid, hemione, asinine, and zebrine), we achieve a completely new pattern of early evolution of Equus. The clade consisting of E. teilhardi and E. yunnanensis forms sister groups with all extant species, distinct from the clade that consists of all other stenonid species. Based on this result, probably the “stenonids” are not a phylogenetic unit any more, but just a morphologic concept. E. teilhardi and E. yunnanensis are not typical stenonid species, they both have relatively short facial part and robust limbs, which are more similar to most extant species. Probably these two species are closer to extant forms rather than other stenonid species.
The EQUUS Datum in China and the Lower Boundary of the Quaternary
Deng and Xue (1997) demonstrated once again the significance of the Equus Datum. They argued that the first appearance of Equus can be considered an easily recognized sign of the lower boundary of the Quaternary in Eurasia. Indeed, the first appearance of Equus was roughly contemporary with many important geological events around 2.5 Ma, such as the Matsuyama/Gauss boundary as determined by magnetostratigraphy, the beginning of loess accumulation in China, the appearance of the ice cap in the Arctic (Backman, 1979) and the first strengthening of neotectonism (Zhu et al., 1994). In recent decades, we put substantial effort into describing the classical localities of the Early Pleistocene in China to correlate the Quaternary mammalian paleontology in China relative to that throughout Eurasia.
Nihewan Basin
In 1924, Barbour observed the strata of Nihewan, Yangyuan, Hebei Province (Figure 2) and established a stratigraphic unit, the Nihewan Beds. Afterwards, Barbour, Licent, Teilhard de Chardin, and Piveteau further studied the Nihewan Beds and their mammalian fossils. In 1948, IGC (International Geology Congress) regarded the Villafranchian as the lower boundary of the terrestrial Pleistocene in Europe, which suggested that the Nihewan Beds represented the lower boundary of the Pleistocene in China. The suggestion was formally approved and used by Chinese geologists in 1954.
Along with research progress, a lot of different viewpoints about the Nihewan Beds have emerged, the main three of which are: (1) the Nihewan Beds are assigned to the Lower Pleistocene; (2) the Nihewan Beds consist of the Lower Pleistocene and Middle Pleistocene; and (3) the Nihewan Beds may include Lower–Middle Pleistocene and Upper Pleistocene, and even Pliocene (Wei and Xie, 1989).
Tang and Ji (1983) established the Dongyaozitou Formation and its accompanying Dongyaozitou Fauna on the basis of the Dongyaozitou section in Yuxian, Hebei Province. Based on the components of this fauna, Deng and Xue (1999a) assigned it to the Pliocene.
Li (1984) discovered typical Nihewan Fauna in the Danangou section above the Dongyaozitou Formation in Yuxian. Most of the fossils are the main members of the Nihewan Fauna. Based on the primitive features of the micromammals, this section was shown to represent the earliest horizon of the Pleistocene (i.e., the Nihewan Formation) (Zheng, 1981). Because the Nihewan Fauna is located between the two formations in Danangou, Yuxian, the first appearance of Equus corresponds to their boundary.
Wu et al. (1995) studied the age of the Nihewan Beds, especially the lower boundary of the Quaternary, by amino acid dating. The dating samples are bones collected from a grayish-white gravel bed in the lower part of the Dongyaozitou Formation in Yuxian, which is 2.78 Ma old. Wu et al. thought that the lower boundary of the Quaternary in this region should be the lower boundary of the Nihewan Formation but above the Dongyaozitou Formation in the Danangou section, and that it was about 2.5 Ma old.
Cai et al. (2004) chose the Danangou section as the type section of Nihewan Formation. They divided this section into four informal lithological members. They noted that Equus was accompanied by hipparionines in member 2. They suggested that the base of member 2 corresponded to the lower boundary of the Quaternary (about 2.58 Ma) based on the micromammalian fossil assemblage. Recently, an increasing number of studies on magnetostratigraphy suggested that the oldest sediments of the Nihewan beds date back to about 2.5 Ma, which is roughly corresponds to the Plio-Pleistocene boundary (Zhu et al., 2007; Deng et al., 2008; Gibbard and Head, 2010; Liu et al., 2012, 2018; Ao et al., 2013a).
Yuanmou Basin
The Yuanmou Basin in Yunnan Province has been the type region of the Early Pleistocene strata in South China (Figure 2), and it has been researched for as long as the Nihewan region. In the 1920s−1930s, Walter Granger collected mammalian fossils from the Yuanmou Basin. In the 1940s, Colbert (1940) studied these fossils and named a fossil horse among them E. yunnanensis. He thought that these fossils corresponded to the fauna of the Upper Irrawaddy in Myanmar and the Early Pleistocene Nihewan Beds in North China based on the horse's small size and some primitive features of its teeth.
Pei (1961) suggested that the Early Pleistocene strata, which had yielded E. yunnanensis, should correspond to the Pinjor and Tatrot Beds of the Upper Siwalik in the Indo-Pakistani subcontinent, and its geological age should correspond to the Villafranchian in Europe. After discovering human fossils in 1965, a lot of researchers began to study this region and proposed some different viewpoints.
The fossiliferous beds in the Yuanmou Basin are included in the Yuanmou Formation, which is subdivided into four members. Li (1981) thought the fossils that were collected from the Yuanmou Basin could be divided into two groups: one group at Members 1–2, which contains Late Pliocene forest mammals, and another group at Members 3–4, which contains some surviving Neogene mammals. The latter's extinction rate was rather high, but there are also some Quaternary mammals that were widespread in North and South China, such as E. yunnanensis. Members 3–4 do not contain forms of the Ailuropoda-Stegodon Fauna; therefore, it should be assigned to the Early Pleistocene.
Qian and Zhou (1991) separated the fossils from Members 1–2 of the Yuanmou Formation from the previously confused “Yuanmou Fauna” and named them the Shagou Fauna. Chow (1961) first thought that this fauna corresponded to that of the Dhok Pathan Beds of the Middle Siwalik in India, which have yielded a similar fauna and date to the Late Pliocene. The Shagou Fauna does not contain Equus. The Xiaohe Fauna is under the Shagou Fauna. Qian et al. thought that the Xiaohe Fauna corresponded to the Middle Siwalik Fauna in South Asia, the Baode Fauna in North China, and the Lufeng Fauna in Yunnan, so it should undoubtedly be assigned to the Pliocene. Qian et al. had a different viewpoint from Chow (1961), but Zhang et al. (1978) and some other authors thought that the Xiaohe Fauna is actually included in the Shagou Formation and this formation belongs to the Pliocene.
The Yuanmou Fauna is located above the Shagou Fauna and collected from Members 3–4 of the Yuanmou Formation. The Yuanmou Fauna contains rich mammalian fossils, including Homo erectus yuanmouensis, which is famous in Chinese paleoanthropology research, in its upper part, mammalian survivors of the Neogene, of which the extinction rate was 93%, and Early Pleistocene mammals. Deng and Xue (1999a) indicated that firstly Yuanmou Fauna contains Early Pleistocene mammal such as E. yunnanensis, Canis yuanmouensis, Hyaena licenti, Stegodon elephantoides, Axis shansius, Axis cf. rugosus, Rusa yunnanensis, etc.; there are also some survivors of the Neogene mammals; and the members which commonly occur in the Ailuropoda-Stegodon fauna in the Pleistocene in south China such as Ailuropoda, Tapirus, Ursus, Pongo, and Macaca. They concluded that Yuanmou Fauna has been shown to be the earliest Early Pleistocene fauna.
Because Equus is absent from the Shagou Fauna, E. yunnanensis is the first appearance of Equus in the Yuanmou Fauna and its earliest appearance was in Member 3 of the Yuanmou Formation. Paleomagnetic dating revealed that the Matsuyama/Gauss boundary, which is located on the bottom of Member 3 (Qian and Zhou, 1991), dates to about 2.58 Ma. Moreover, according to the fission track dating performed by Qian and Zhou (1991), the age of the upper section of Shagou Formation was 2.59 Ma. Equus yunnanensis happens to appear at this boundary, which thus corresponds well with the lower boundary of the Quaternary.
According to a highly resolved magnetostratigraphic analysis performed by Zhu et al. (2008), the sediments of the Yuanmou Basin, which were divided into 17 polarity units, should be grouped into three distinct directional intervals. The base of Group I, which represented the lower boundary of the Quaternary, dates to 2.58 Ma.
Linxia Basin
The Linxia Basin is located in the east of Gansu Province (Figure 2). Since the end of the 1980s, the Linxia Basin has been famous for its wealth of fossil mammals. In May of 1999, while identifying the “dragon bones” obtained by a local “dragon bone” dealer, Zhanxiang Qiu noticed some canid fossils from loess deposits. Based on the dealer's information, Qiu found a very fossiliferous locality in Longdan Village, Dongxiang Autonomous County. After about 1 year, Qiu and his colleagues collected more than 100 specimens, mainly skulls and mandibles. Consequently, the Longdan Fauna became one of the most important Early Pleistocene mammalian faunas in China.
The Longdan Fauna described by Qiu et al. (2004) includes 31 species of 29 genera. The presence of Equus and the absence of Middle Pleistocene species indicates that the age of the Longdan Fauna is Early Pleistocene. Additionally, Wang (2005), Qiu et al. (2009), and Deng (2012), respectively, reported specimens of Castor anderssoni, Ursus yinanensis, and Proboscidipparion sinense from Longdan. Qiu et al. noted that the Longdan Fauna and the Nihewan Fauna shared only eight species. They determined that the Simpson similarity index (i.e., the number of common taxa divided by the number of total taxa of the fauna with fewer taxa) was only 25.8%. The number of genera commonly shared by the two faunas is 17, and the similarity index at the generic level is thus 58.6%.
Qiu et al. (2004) suggested that differences in composition between the Longdan and Nihewan Faunas may reflect differences in geologic ages and paleoenvironments. The Longdan Fauna was slightly older than the Nihewan Fauna based on the observation that the Longdan Fauna contains some primitive forms and the Nihewan Fauna includes a large number of advanced forms. For example, the extremely large horse E. eisenmannae represented a very primitive lineage that was similar to the earliest horse in Europe, E. livenzovensis.
Although no agreement has been reached regarding the geologic age of the Nihewan Fauna, many paleontologists consider it to be around 1.8 Ma (Liu and Liu, 2000; Qiu, 2000). Previous paleomagnetic data indicated that the Longdan is 2.55–2.16 Ma old (Qiu et al., 2004). The results of paleoenvironmental and chronological analysis performed by Zan et al. (2016) demonstrated that the age of the Longdan Fauna was 2.5–2.2 Ma.
Other Sections
The loess in southeastern Shanxi Province is, from bottom to top, divided into Old Red Loam, New Red Loam, Lishi Loess, Malan Loess, and Holocene Loess. The Old Red Loam is distributed in the eastern Yushe Basin. Cao et al. (1995) discovered a lot of mammalian fossils in the basal conglomerate bed of the Old Red Loam. Most of these fossils are teeth of Equus sp., and they are intact and not abraded, which means that they are probably not reworked and most likely represent the true horizon at which Equus appears. Under the Old Red Loam, the Yushe Group contains hipparionines but not Equus. According to paleomagnetic studies (Shi, 1994; Shi et al., 1994), the Old Red Loam was deposited 2.5–1.5 Ma; therefore, its lower boundary was 2.5 Ma. The Equus fossils were collected from the basal conglomerate bed of this boundary.
In the Youhe District in Weinan, Shaanxi Province (Figure 2), the strata that contained the Early Pleistocene Nihewan Fauna are yellowish-brown coarse deposits, and they are informally called “Yellow Sanmen”; the strata that contained the Pliocene Youhe Fauna are green or grayish-green mudstones or marls, and are called “Green Sanmen.” In the Youhe section, Equus first appears in the Yellow Sanmen, under which the Green Sanmen is only observed from Borehole W7 in Yancun, Weinan, which indicates that the Matsuyama Chron is recorded in the Yellow Sanmen, which dates to approximately 3.0–2.58 Ma. Zhu et al. (1995) conducted a paleomagnetic study on the Songjiabeigou section in Youhe, Weinan, and they also found that the Matsuyama/Gauss boundary is located between the Sanmen Formation (i.e., the Yellow Sanmen) and the Youhe Formation (i.e., the Green Sanmen). Consequently, the sections in the Youhe District demonstrate that the first appearance of Equus was at the Matsuyama/Gauss boundary at 2.58 Ma. Tang et al. (1983) described the Linyi Fauna, which included E. huanghoensis reported by Chow and Chow (1965). Based on the composition, Tang et al. (1983) stated that the age of the Linyi Fauna corresponded to the middle to late Villafranchian, which is the same as the Sanmen Formation. Therefore, the first occurrence of E. huanghoensis should be about 2.58 Ma at the lower boundary of the Quaternary.
In 1961, Xue and Wang collected plenty of mammalian fossils from Bajiazui, Qingyang, Gansu Province (Deng and Xue, 1999a; Figure 2). These fossils were obtained from sandstone at the base of the section, the age of which was identified as Early Pleistocene (Wang et al., 1966; Wang and Xue, 1982). Deng and Xue (1999a,b,c) studied the Equus specimens of this mammalian fauna and erected two new species, E. qingyangensis and E. wangi, and identified a known species, E. teilhardi. Deng and Xue (1999a) indicated that E. qingyangensis occurred around 2.5 Ma based on the position of fossils and loess between fluvial-lacustrine deposits. As we discussed above, E. qingyangensis was very primitive, especially on its very low cranial proportions, which are similar to those of the earliest Equus in the world, E. shoshonensis (Figure 12).
Scenario of EQUUS Datum and Early Evolution
Based on the information detailed above, we can describe the pattern of the early evolution of stenonids in China. There is no doubt that stenonids first arrived in China at the lower boundary of the Quaternary. It also can be confirmed that at least five stenonid species simultaneously appeared in China about 2.5 Ma (Figure 14). This is not strange. Equids are all highly cursorial and able to migrate very long distances in a relatively short time. When stenonids arrived in Eurasia and dispersed, the time difference of their dispersal was negligible on a geochronological scale. But why could stenonids radiate and produce high diversity as soon as they appeared in Eurasia, even only in China? Was that a coincidence?
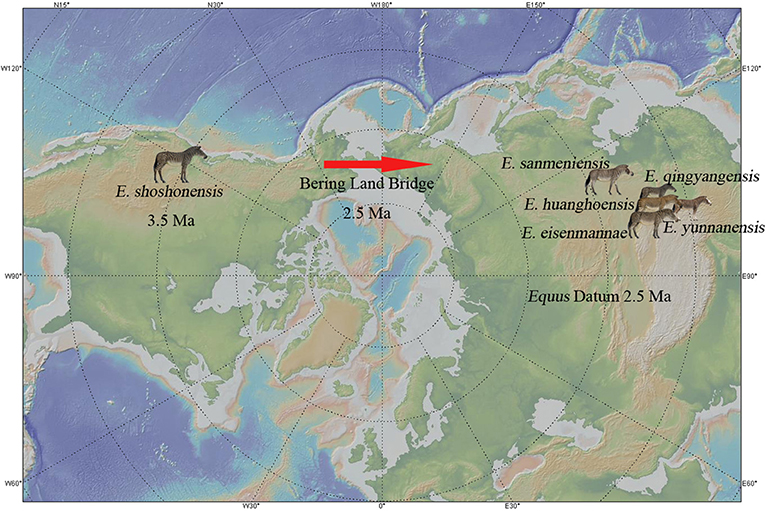
Figure 14. Origin of Equus in North America and the Equus Datum in China.
Equidae originated in North America 56 Ma and was restricted to this continent from the Eocene to the Oligocene (MacFadden, 2005; Bai et al., 2018). Since the Early Miocene, there have been several migrations of Equidae to the Old World (MacFadden, 2005). This is probably related to the Mi-1 glaciation (Zachos et al., 2001). Based on diversity, geographic distribution, and population, Anchitheriinae was not a prosperous lineage in Eurasia. For example, only two genera with four species of Anchitheriinae have been found in China (Hou et al., 2007). In the beginning of the Late Miocene, a much more successful clade, Hipparionini, appeared in Eurasia. The earliest species was Hipparion dongxiangense, which was found in Guonigou, Dongxiang, Gansu Province; the first occurrence of H. dongxiangense was 11.5 Ma (Fang et al., 2016). Throughout the Late Miocene, hipparionine originated and diversified into a successful taxon in China, with 8 genera and 17 species. In the Pliocene, four genera with three species of hipparionine survived, and one new genus and three new species originated (Qiu et al., 1987; Li and Li, 1990; Sun, 2013; Deng et al., 2016; Bernor et al., 2018; Sun et al., 2018; Figure 13). Although the diversity decreased significantly, Hipparionini was still the main member of the mammalian fauna in the Pliocene. In the Pliocene, the first appearance of Equus was in North America (3.5 Ma, Winans, 1989). According to O'Regan et al. (2010), the Bering Strait was likely exposed about 3.3 Ma, which allowed the North American Equus species to invade Eurasia (Figure 15). However, they likely did not migrate until later because hipparionine species were still the dominant equids in Eurasia, at least in East Asia (Figure 13).
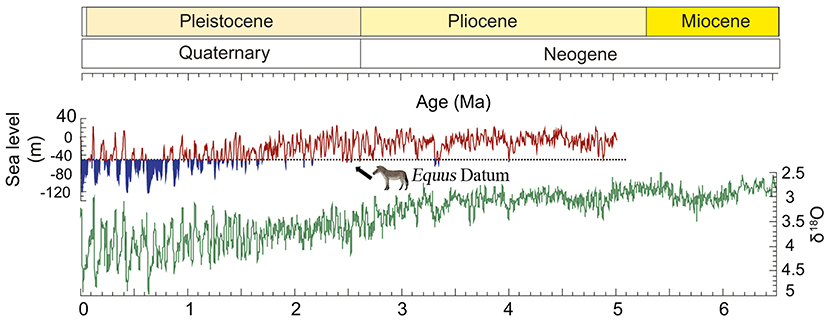
Figure 15. Equus Datum, global sea-level and global benthic δ18O compilation. Blue shading of the sea-level curve indicates times when global sea-level dropped below 50 m and the Bering Strait was likely exposed (modified after O'Regan et al., 2010).
Stable carbon isotopic analysis performed by Hou et al. (2006) indicated that the habitats in western China were steppes dominated by C3 grasses during the Late Miocene and Early Pliocene instead of savannas dominated by C4 plants. They suggested that C4 plants dispersed into northern China in the Pliocene along with the strengthening of the East Asian summer monsoon. Simultaneously, the diversity of the Hipparion fauna in East Asia gradually declined. They also suggested that Miocene equids in North America faced a similar crisis because of C4 plant dispersal 7–5 Ma. The dispersal of C4 plants has been explained to be correlated with a decrease in atmospheric CO2 (Cerling et al., 1993, 1997; Wang et al., 1994). Janis et al. (2000) suggested that the observed decline in species richness of North American horses may represent a gradual decline in primary productivity, which would be consistent with a decrease in atmospheric CO2. In summary, we propose that, after the Early Pliocene, hipparionine species declined as a result of decreased primary productivity of vegetation in their habitats.
The massive recession of Hipparion was initiated in the Early Pleistocene, by which only two genera and two species survived (Qiu et al., 1987; Deng, 2012; Bernor et al., 2015). This recession was mostly the result of accumulation and aggravation of the constant decline of primary productivity of the vegetation and partly influenced by climate change. The beginning of the Pleistocene was when Equus first immigrated into Eurasia. As described in our discussion above, at least five stenonid species (E. qingyangensis, E. eisenmannae, E. sanmeniensis, E. huanghoensis, and E. yunnanensis) simultaneously appeared in China at the very beginning of the Pleistocene. Compared with hipparionine, stenonids had obvious advantages. First, monodactyl feet adapted to vertical standing made Equus the perfect steppe runner. In addition, the V-scars of Equus are developed on the first phalanx of both fore and hind digit III, and are much wider and flatter than those of hipparionines (Deng et al., 2012). Additionally, all stenonid species have very elongated face. Qiu et al. (2004) indicated that the elongated jaw could accommodate a larger cheek tooth row, which would be a powerful adaptation for chewing coarse food, like dry grass. In contrast, hipparionines in the Early Pleistocene, such as Proboscidipparion sinense, had relatively small cheek teeth. Moreover, the global climate became colder in the Early Pleistocene, and the animals with larger body sizes were better adapted to the cold environment according to Bergmann's rule (Qiu et al., 2004). The famous Early Pleistocene fauna, the Nihewan Fauna, consisted of large animals, such as, Coelodonta nihowanensis, Elasmotherium sp., Paracamelus gigas, and Bison palaeosinensis (Cai et al., 2013). The stenonid horses, especially E. eisenmannae, are very large. Only very large hipparionine species, including Proboscidipparion sinense and Plesiohipparion shanxiense, survived in the beginning of the Pleistocene in China (Qiu et al., 1987; Deng, 2012; Bernor et al., 2015); consequently, plenty of open habitats and ecological niches were available for the new incoming lineages. As Equus arrived in Eurasia, its adaptations to this new situation helped it radiate and disperse rapidly. Deng et al. (1999, 2002) performed carbon isotopic analysis on mammalian fossils in the Quaternary in China and revealed an environment dominated by C3 plants in northern China in the Early Pleistocene that was led by strengthening of the East Asian winter monsoon along with Quaternary glaciation. Therefore, the primary productivity of vegetation likely increased significantly, and stenonid horses, which had been widespread in northern China, faced an opportunity for further evolution.
Equus shoshonensis has often been regarded as ancestral to all stenonids and even all Equus species (Deng and Xue, 1999a). Skinner and Hibbard (1972) argued that there were marked similarities between the skulls and dentitions of the extinct North American E. shoshonensis (his E. simplicidens) and the living Grévy's zebra, E. grevyi. Based on our observation and comparison of E. shoshonensis fossils and specimens of extant E. grevyi housed in several museums, these two species are similar in absolute size, and in cranial and distal limb proportions. However, our phylogenetic analysis places E. grevyi in the smallest clade formed by extant horses (Figures 1, 13). The similarities discussed above, which are referred to functional morphology, should relate to similar habitat. E. grevyi lives in a semi-desert environment in northern East Africa with negative mean annual climatic water balance (Schulz and Kaiser, 2013). Equus shoshonensis probably lived in a similar environment to that of modern E. grevyi. Therefore, Equus was probably adapted to arid environments from the very beginning. When the Bering land bridge opened at the beginning of the Pleistocene (Figures 14, 15), Equus immigrated into Eurasia from North America, where it first stopped in East Asia. Since 2.5 Ma, a stronger winter monsoon system and higher continental desiccation occurred in China, most probably due to strong uplift of the Tibetan Plateau (An et al., 2001). Equus was well-adapted to this climate and rapidly radiated into a considerably dominant member of the Early Pleistocene mammalian fauna, the Equus Fauna.
Data Availability Statement
All datasets for this study are included in the article/Supplementary Material.
Author Contributions
BS and TD designed the conceptual model and contributed equally to the writing of the manuscript.
Funding
This work was supported by the Chinese Academy of Sciences (grant nos. XDB26030302, XDA20070203, QYZDY–SSW–DQC022, and GJHZ1885), the Special Research Program of Basic Science and Technology of the Ministry of Science and Technology (grant no. 2015FY310100), and the National Natural Science Foundation of China (41430102).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The editor Dr. Leonardo Santos Avilla, reviewers Dr. David Marjanović and Dr. Xiaoke Qiang greatly improve our manuscript. Mr. Yu Chen (IVPP) provided art work in our manuscript. Dr. Shiqi Wang (IVPP) and Mr. Hanwen Zhang (University of Bristol) provided pictures of skull specimens. Dr. Wenhui Liu (National Museum of China) provided very important literatures for references. Drs. Haibing Wang and Shunxing Jiang (IVPP) provided helpful support on phylogenetic work. Dr. Guillaume Billet (Muséum national d'Histoire naturelle) provided access to observations on type specimens of the Nihewan fauna. Mses. Judy Galkin and Eleanor Hoeger (American Museum of Natural History) respectively provided accesses to observations on specimens of fossil horses and extant zebras. We appreciate all of their contributions.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00429/full#supplementary-material
Abbreviations
IGF, Instituto geologico di Firenze, Florence, Italy; IVPP V, vertebrate paleontology collection of Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; NHMUK PV M, cast at the Natural History Museum, London, UK; NIH, Nihewan collection of the Muséum national d'Histoire naturelle, Paris, France; NWUV, Institute of Cenozoic Geology and Environment, Northwest University, Xi'an, China; PMU M, Museum of Evolution of Uppsala University, Uppsala, Sweden; THP, Tianjin Natural History Museum, Tianjin, China.
References
Alberdi, M. T., Ortiz-Jaureguizar, E., and Prado, J. L. (1998). A quantitative review of European stenonid horses. J. Paleontol. 72, 371–387. doi: 10.1017/S0022336000036350
An, Z., Kutzbach, J. E., Prell, W. L., and Poter, S. C. (2001). Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan Plateau since late Miocene times. Nature 411, 62–66. doi: 10.1038/35075035
Ao, H., An, Z., Dekkers, M. J., Li, Y., Xiao, G., Zhao, H., et al. (2013a). Pleistocene magnetochronology of the fauna and Paleolithic sites in the Nihewan Basin: Significance for environmental and hominin evolution in North China. Q. Geochronol. 18, 78–92. doi: 10.1016/j.quageo.2013.06.004
Ao, H., Dekkers, M. J., An, Z., Xiao, G., Li, Y., Zhao, H., et al. (2013b). Magnetostratigraphic evidence of a mid-Pliocene onset of the Nihewan Formation-implications for early fauna and hominid occupations in the Nihewan Basin, North China. Q. Sci. Rev. 59, 30–42. doi: 10.1016/j.quascirev.2012.10.025
Azzaroli, A. (1965). On the two Villafranchian horses of the Upper Valdarno. Palaeont. Ital. 59, 1–12.
Azzaroli, A. (1982). On Villafranchian palaeoarctic equids and their allies. Palaeont. Ital. 72, 74–97.
Azzaroli, A. (1987). On the occurrence of Equus stenonis in China. Boll. Soc. Paleontol. Ital. 25, 199–201.
Azzaroli, A. (1992). Ascent and decline of monodactyl equids: a case for prehistoric overkill. Ann. Zool. Fennici 28, 151–163.
Azzaroli, A., de Giuli, C., Ficcarelli, G., and Torre, D. (1988). Late Pliocene to early mid-Pleistocene mammalian faunas in Eurasia: faunal succession and dispersal events. Palaeogeogr. Palaeoclimatl. Palaeoecol. 66, 77–100. doi: 10.1016/0031-0182(88)90082-x
Azzaroli, A., and Voorhies, M. R. (1993). The genus Equus in North America, the Blancan species. Palaeont. Ital. 80, 175–198.
Backman, T. (1979). Pliocene biostratigraphy of DSDP 111 and 116 from the northern Atlantic Ocean and of northern hemisphere glaciations. Stockholm Contr. Geol. 33, 115–137.
Bai, B., Wang, Y., and Meng, J. (2018). The divergence and dispersal of early perissodactyls as evidenced by early Eocene equids from Asia. Commun. Biol. 1:115. doi: 10.1038/s42003-018-0116-5
Bennett, D. K. (1980). Stripes do not a zebra make, part I: a cladistic analysis of Equus. Syst. Zool. 29, 272–287. doi: 10.2307/2412662
Bernor, R. L., Sun, B., and Chen, Y. (2015). Plesiohipparion shanxiense n. sp. from the Early Pleistocene (Nihowanian) of Shanxi, China. Boll. Soc. Paleontol. Ital. 54, 197–210.
Bernor, R. L., Wang, S., Liu, Y., Chen, Y., and Sun, B. (2018). Shanxihippus dermatorhinus (new gen.) with comparisons to Old World hipparions with specialized nasal apparati. Riv. Ital. Paleontol. Stratigr. 124, 361–386.
Cai, B., Zhang, Z., Zheng, S., Qiu, Z., Li, Q., and Li, Q. (2004). New advances in the stratigraphic study on representative sections in the Nihewan Basin, Hebei. Prof. Pap. Stratigr. Paleont. 28, 267–285.
Cai, B., Zheng, S., Liddicoat, J. C., and Li, Q. (2013). “Review of the Litho-, Bio-, and chronostratigraphy in the Nihewan Basin, Hebei, China,” in Fossil Mammals of Asia, Neogene Biostratigraphy and Chronology, eds X. Wang, L. J. Flynn, and M. Fortelius (New York, NY: Columbia University Press), 218–242. doi: 10.7312/columbia/9780231150125.003.0008
Cao, J., Shi, N., and Zhang, J. (1995). A study of loess in southeastern Shanxi, China. Quat. Sci. 123–129.
Cerling, T. E., Harris, J., MacFadden, B. J., Leakey, M. G., Quade, J., Eisenmann, V., et al. (1997). Global vegetation change through the Miocene/Pliocene boundary. Nature 389, 153–158. doi: 10.1038/38229
Cerling, T. E., Wang, Y., and Quade, J. (1993). Global ecological change in the late Miocene: expansion of C4 ecosystems. Nature 361, 344–345. doi: 10.1038/361344a0
Chow, M., and Chow, B. (1965). Notes on Villafranchian mammals of Lingyi, Shansi. Vert. PalAsiat. 9, 223–234.
Colbert, E. (1940). Pleistocene mammals from the Ma Kai Valley of northern Yunnan, China. Am. Mus. Novitates 1099, 1–10.
Dalquest, W. W. (1988). Astrohippus and the origin of Blancan and Pleistocene horses. Occ. Pap. Mus. Texas Technical Univ. 116, 1–23. doi: 10.5962/bhl.title.156562
Deng, C., Zhu, R., Zhang, R., Ao, H., and Pan, Y. (2008). Timing of the Nihewan formation and faunas. Quat. Res. 69, 77–90. doi: 10.1016/j.yqres.2007.10.006
Deng, T. (2012). A skull of Hipparion (Proboscidipparion) sinense (Perissodactyla, Equidae) from Longdan, Dongxiang of northwestern China – addition to the Early Pleistocene Longdan mammalian fauna (3). Vert. PalAsiat. 50, 74–84.
Deng, T., Dong, J., and Wang, Y. (2002). Variation of terrestrial ecosystem recorded by stable carbon isotopes of fossils in northern China during the Quaternary. Chin. Sci. Bull. 47, 76–78. doi: 10.1360/02tb9015
Deng, T., Li, Q., Tseng, Z. J., Takeuchi, G. T., Wang, Y., Xie, G. P., et al. (2012). Locomotive implication of a Pliocene three-toed horse skeleton from Tibet and its paleo-altimetry significance. Proc. Natl. Acad. Sci. U.S.A. 109, 7374–7378. doi: 10.1073/pnas.1201052109
Deng, T., Wang, H., Wang, X., Li, Q., and Tseng, Z. J. (2016). The Late Miocene Hipparion (Equidae, Perissodactyla) fossils from Baogeda Ula, Inner Mongolia, China. Histor. Biol. 28, 53–68. doi: 10.1080/08912963.2015.1020425
Deng, T., and Xue, X. (1997). Redemonstrating the first appearance of the genus Equus as a sign of the lower boundary of the Quaternary. J. Stratigr. 21, 109–116
Deng, T., and Xue, X. (1999a). Chinese Fossil Horses of Equus and Their Environment. Beijing: China Ocean Press.
Deng, T., and Xue, X. (1999b). Equus qingyangensis sp. nov. (Perissodactyla, Equidae) from the Early Pleistocene of Qingyang, Gansu, China. Vert. PalAsiat. 37, 62–74.
Deng, T., and Xue, X. (1999c). “A new species of stenonid horse from China,” in Proceedings of the Seventh Annual Meeting of the Chinese Society of Vertebrate Paleontology, eds Y. Wang and T. Deng (Beijing: China Ocean Press, 189–200.
Deng, T., and Xue, X. (1999d). Phylogenetic relationships of the Chinese fossil species of the genus Equus (Perissodactyla, Equidae). Sci. China 42, 449–455. doi: 10.1007/BF02875238
Deng, T., Xue, X., and Dong, J. (1999). The evidence of fossil carbon isotopes of the climatic event at the beginning of Quaternary. Chin. Sci. Bull. 44, 477–480. doi: 10.1007/BF02977893
Dong, W., and Fang, Y. S. (2005). Fossil equids (mammals) from the Tuozidong, Nanjing (China) and its significance. Vert. PalAsiat. 43, 36–48.
Eisenmann, V. (1975). Nouvelles interpretations des restes d'équidés (Mammalia, Perissodactyla) de Nihowan (Pléistocène inférieur de la Chine du Nord): Equus teilhardi nov. sp. Geobios 8, 125–134. doi: 10.1016/S0016-6995(75)80009-X
Eisenmann, V. (1996). “Quaternary horses: possible candidates to domestication,” in Proceedings of the XIII Congress (Forlì: International Union of Prehistoric and Protohistoric Sciences), 8–14.
Eisenmann, V., and Baylac, M. (2000). Extant and fossil Equus (Mammalia, Perissodactyla) skulls: a morphometric definition of the subgenus Equus. Zool. Scr. 29, 89–100. doi: 10.1046/j.1463-6409.2000.00034.x
Eisenmann, V., and Deng, T. (2005). Equus qingyangensis (Equidae, Perissodactyla) of the Upper Pliocene of Bajiazui, China: evidence for the North American origin of an Old World lineage distinct from E. stenonis. Quaternaire 2, 113–122.
Eisenmann, V., and Karchoud, A. (1982). Analyses multidimension nelles de métapodes d'Equus sensu lato (Mammalia, Perissodactyla). Bulletin du Muséum National d'Histoire naturelle, Section C: Sciences de la Terre – Paléontologie, Geologie, Minéralogie 4, 75–103.
Fang, X., Wang, J., Zhang, W., Zan, J., Song, C., Yan, M., et al. (2016). Tectonosedimentary evolution model of an intracontinental flexural (foreland) basin for paleoclimatic research. Glob. Planetary Change 145, 78–97. doi: 10.1016/j.gloplacha.2016.08.015
Forsten, A. (1984). Supraspecific grouping of Old World hipparions (Mammalia, Equidae). Paläontol. Z. 58, 165–171. doi: 10.1007/BF02990336
Forsten, A. (1988). The small caballoid horse of the upper Pleistocene and Holocene. J. Anim. Breed. Genet. 105, 161–176. doi: 10.1111/j.1439-0388.1988.tb00288.x
Gibbard, P. L., and Head, M. J. (2010). The newly-ratified definition of the Quaternary System/Period and redefinition of the Pleistocene Series/Epoch, and comparison of proposals advanced prior to formal ratification. Episodes 33, 152–158. doi: 10.18814/epiiugs/2010/v33i3/002
Goloboff, P. A., Farris, J. S., and Nixon, K. C. (2008). TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786. doi: 10.1111/j.1096-0031.2008.00217.x
Gray, J. E. (1821). On the natural arrangement of vertebrose animals. Lond. Med. Repos. 15, 296–310.
Hopwood, A. T. (1936). The former distribution of caballine and zebrine horses in Europe and Asia. Proc. Zool. Soc. Lond. 106, 897–912. doi: 10.1111/j.1469-7998.1936.tb06291.x
Hou, S., Deng, T., He, W., and Chen, S. (2007). New materials of Sinohippus from Gansu and Nei Mongol, China. Vert. PalAsiat. 45, 213–231.
Hou, S., Deng, T., and Wang, Y. (2006). “Stable carbon isotopic evidence of tooth enamel for the late Neogene habitats of the Hipparion fauna in China,” in Proceedings of the Tenth Annual Meeting of the Chinese Society of Vertebrate Paleontology, ed W. Dong (Beijing: China Ocean Press, 85–94.
Janis, C. M., Damuth, J., and Theodor, J. M. (2000). Miocene ungulates and terrestrial primary productivity: where have all the browsers gone? Proc. Natl. Acad. Sci. U.S.A. 97, 7899–7904. doi: 10.1073/pnas.97.14.7899
Li, F., and Li, D. (1990). “Latest Miocene Hipparion (Plesiohipparion) of Zanda Basin,” in Palaeontology of the Ngari Area, Tibet (Xizang), eds Z. Yang and Z. Nie (Wuhan: China University of Geoscience Press, 186–193.
Li, Y. (1981). On the subdivisions and evolution of the Quaternary mammalian faunas of South China. Vert. PalAsiat. 19, 67–76.
Li, Y. (1984). The early Pleistocene mammalian fossils of Danangou, Yuxian, Hebei. Vert. PalAsiat. 22, 60–68.
Li, Y., Zhang, Y., Sun, B., Ao, H., and Xue, X. (2016). New fossils of the Early Pleistocene Equus huanghoensis (Equidae, Perissodactyla) from Nihewan in Hebei province of China. Sci. China Earth Sci. 59, 83–94. doi: 10.1007/s11430-015-5138-y
Linnaeus, C. (1758). Systema Naturae, Per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species cum Characteribus, Differentiis, Synonymis, Locis. Editio Decimal. Stockholmiae: Salvii. doi: 10.5962/bhl.title.542
Liu, H., and You, Y. (1974). New Materials of Equus yunnanensis in Yuanmou, Yunnan. Vert. PalAsiat. 12, 126–136.
Liu, J., and Liu, Q. (2000). Quaternary stratigraphy in China. Quat. Sci. 20, 129–141. doi: 10.3321/j.issn:1001-7410.2000.02.003
Liu, P., Deng, C., Li, S., Cai, S., Cheng, H., Yuan, B., et al. (2012). Magnetostratigraphic dating of the Xiashagou fauna and implication for sequencing the mammalian faunas in the Nihewan Basin, North China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 315–316, 75–85. doi: 10.1016/j.palaeo.2011.11.011
Liu, P., Yue, F., Liu, J., Qin, H., Li, S., Zhao, X., et al. (2018). Magnetostratigraphic dating of the Shixia red sediments and implications for formation of Nihewan paleo-lake, North China. Quat. Sci. Rev. 193, 118–128. doi: 10.1016/j.quascirev.2018.06.013
MacFadden, B. J. (2005). Fossil horses – evidence for evolution. Science 307, 1728–1730. doi: 10.1126/science.1105458
MacFadden, B. J. (2006). “Early Pliocene (latest Hemphillian) horses from the Yepómera Local 33 Fauna, Chihuahua, Mexico,” in Advances in Late Tertiary Vertebrate Paleontology in Mexico and the Great American Biotic Interchange, eds Ó. Carranza-Castañeda and E. H. Lindsay (México: Universidad Nacional Autónoma de México, Instituto de Geología and Centro de Geociencias, Publicación Especial 4, 33–43.
Matsumoto, H. (1915). On some fossil mammals from Honan, China. Sci. Rep. Tohoku Univ. Ser. 3, 29–38.
O'Regan, M., John, K. S., and Moran, K. (2010). Plio-Pleistocene trends in ice rafted debris on the Lomonosov Ridge. Quat. Int. 219, 168–176. doi: 10.1016/j.quaint.2009.08.010
Owen, R. (1848). Description of the teeth and portions of jaws of two extinct anthracotheroid quadrupeds (Hyopotamus vectianus and Hyop. bovinus) discovered by the Marchioness of Hastings in the Eocene deposits of the N.W. coast of the Isle of Wight: with an attempt to develop Cuvier's idea of the classification of the pachyderms by the number of their toes. Q. J. Geol Soc. 4, 103–141. doi: 10.1144/GSL.JGS.1848.004.01-02.21
Pagnac, D. (2006). Scaphohippus, a new genus of horse (Mammalia: Equidae) from the Barstow Formation of California. J. Mammal. Evol. 13, 37–61. doi: 10.1007/s10914-005-9002-2
Pei, W. (1961). Fossil mammals of Early Pleistocene age from Yuanmo (MaKai) of Yunnan. Vert PalAsiat. 1, 16–30.
Prado, J. L., and Alberdi, M. T. (1996). A cladistic analysis of the horses of the tribe Equini. Palaeontology 39, 663–680.
Qian, F., and Zhou, G. (1991). Quaternary Geology and Paleoanthropology of Yuanmou, Yunnan, China. Beijing: Science Press.
Qiu, Z. (2000). Nihewan fauna and Q/N boundary in China. Quat. Sci. 20, 142–154. doi: 10.3321/j.issn:1001-7410.2000.02.004
Qiu, Z., Deng, T., and Wang, B. (2004). Early Pleistocene mammalian fauna from Longdan, Dongxiang, Gansu, China. Palaeont. Sin. New Ser. C 27, 1–198.
Qiu, Z., Deng, T., and Wang, B. (2009). First ursine bear material from Dongxiang, Gansu – addition to the Longdan mammalian fauna (2). Vert. PalAsiat. 47, 245–264.
Qiu, Z., Huang, W., and Guo, Z. (1987). The Chinese hipparion fossils. Palaeont. Sin. New Ser C. 25, 1–250.
Qiu, Z., and Tedford, R. H. (2013). “Chapter 2: History of scientific exploration of Yushe Basin,” in Late Cenozoic Yushe Basin, Shanxi Province, China: Geology and Fossil Mammals, History, Geology, and Magnetostratigraphy, Vol. 1, eds R. H. Tedford, Z. Qiu, and L. J. Flynn (Dordrecht: Springer), 69–78. doi: 10.1007/978-90-481-8714-0_2
Schulz, E., and Kaiser, T. M. (2013). Historical distribution, habitat requirements and feeding ecology of the genus Equus (Perissodactyla). Mammal Rev. 43, 111–123. doi: 10.1111/j.1365-2907.2012.00210.x
Shi, N. (1994). The Late Cenozoic stratigraphy, chronology, palynology and environment development in the Yushe Basin, North China. Striae 36, 1–90.
Shi, N., Liu, H., and Lu, W. (1994). Study of magnetism on Wangning red loam section in Yushe Basin, SE Shanxi Province: its age and sedimentary environment. Quat. Sci. 183–191.
Skinner, M. F., and Hibbard, C. W. (1972). Early Pleistocene pre-glacial and glacial rocks and faunas of North-Central Nebraska. Bull. Am. Mus. Nat. Hist. 148, 117–125.
Sun, B. (2013). The Miocene Hipparion (Equidae, Perissodactyla) from Shihuiba Locality, Lufeng, Yunnan. Vert. PalAsiat. 51, 141–161.
Sun, B., Deng, T., and Liu, Y. (2017). Early Pleistocene Equus (Equidae, Perissodactyla) from Andersson Loc. 32 in Qixian, Shanxi, China. Hist. Biol. 31, 211–222 doi: 10.1080/08912963.2017.1357718
Sun, B., Zhang, X., Liu, Y., and Bernor, R. L. (2018). Sivalhippus ptychodus and Sivalhippus platyodus (Perissodactyla, Mammalia) from the late Miocene of China. Riv. Ital. Paleontol. Stratigr. 124, 1–22. doi: 10.13130/2039-4942/9523
Tang, Y., and Ji, H. (1983). A Pliocene-Pleistocene transitional fauna from Yuxian, northern Hebei. Vert. PalAsiat. 21, 245–254.
Tang, Y., Zong, G., and Xu, Q. (1983). Mammalian fossils and stratigraphy of Linyi, Shanxi. Vert. PalAsiat. 21, 77–86.
Teilhard de Chardin, P., and Piveteau, J. (1930). Les mammifères fossiles de Nihowan (Chine). Ann. Paléont. 19, l−134.
Wang, B. (2005). Beaver (Rodentia, Mammalia) fossils from Longdan, Gansu, China – addition to the Early Pleistocene Longdan mammalian fauna (1). Vert. PalAsiat. 43, 237–242.
Wang, S., and Deng, T. (2011). Some evolutionary trends of Equus eisenmannae (Mammalia, Perissodactyla) in the stratigraphic sequence of Longdan, China, in comparison to modern Equus. J. Vertebr. Paleontol. 31, 1356–1365. doi: 10.1080/02724634.2011.611203
Wang, Y., Cerling, T. E., and MacFadden, B. (1994). Fossil horses and carbon isotopes: new evidence for Cenozoic dietary, habitat, and ecosystem changes in North America. Palaeogeogr. Palaeoclimatol. Palaeoecol. 107, 269–279. doi: 10.1016/0031-0182(94)90099-X
Wang, Y., Hsueh, H., Ho, J., and Chang, K. (1966). Division of the Quaternary strata in the loess district of north Shansi and east Kansu. Acta. Geo. Sin. 46, 102–117.
Wang, Y., and Xue, X. (1982). “Division of loess strata in the middle reaches of the Yellow River,” in Loess & Quaternary Geology (Xi'an: Shaanxi People Press), 1–17
Wei, Q., and Xie, F. (1989). Selected Treatises on Nihewan. Beijing: Cultural Relies Publishing House.
Winans, M. C. (1989). “A quantitative study of the North American fossil species of the genus Equus,” in The Evolution of Perissodactyls, eds D. R. Prothero and R. M. Schoch (New York, NY: Oxford University Press, 262–297.
Wolf, D., Bernor, R. L., and Hussain, T. (2013). A systematic, biostratigraphic, and paleobiogeographic reevaluation of the Siwalik hipparionine assemblage from the Potwar Plateau, northern Pakistan. Palaeontogr. A 300, 1–115. doi: 10.1127/pala/300/2013/1
Wu, P., Long, T., and Wang, S. (1995). Amino acid dating of Nihewan beds. Sci. Geol. Sin. 30, 161–165.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K. (2001). Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292, 686–693. doi: 10.1126/science.1059412
Zan, J., Fang, X., Zhang, W., Yan, M., and Zhang, T. (2016). Palaeoenvironmental and chronological constraints on the Early Pleistocene mammal fauna from loess deposits in the Linxia Basin, NE Tibetan Plateau. Quat. Sci. Rev. 148, 234–242. doi: 10.1016/j.quascirev.2016.07.026
Zhang, Y., You, Y., Ji, H., and Ding, S. (1978). Cenozoic stratigraphy of Yunnan. Prof. Pap. Stratigr. Paleont. 7, 1–21.
Zhu, R., Deng, C., and Pan, Y. (2007). Magnetochronology of the fluvio-lacustrine sequences in the Nihewan Basin and its implications for early human colonization of northeast Asia. Quat. Sci. 27, 922–944. doi: 10.3321/j.issn:1001-7410.2007.06.008
Zhu, R., Potts, R., Pan, Y., Lü, L., Yao, H., Deng, C., et al. (2008). Paleomagnetism of the Yuanmou Basin near the southeastern margin of the Tibetan Plateau and its constraints on late Neogene sedimentation and tectonic rotation. Earth and Planet Sci. Lett. 272, 97–104. doi: 10.1016/j.epsl.2008.04.016
Zhu, R., Yue, L., and Bai, L. (1995). Progress of Quaternary paleomagnetism in China. Quat. Sci. 162–173.
Keywords: Equus Datum, stenonid, evolution, environment change, paleozoogeography
Citation: Sun B and Deng T (2019) The Equus Datum and the Early Radiation of Equus in China. Front. Ecol. Evol. 7:429. doi: 10.3389/fevo.2019.00429
Received: 30 January 2019; Accepted: 23 October 2019;
Published: 13 November 2019.
Edited by:
Leonardo Santos Avilla, Universidade Federal de Estado de Rio de Janeiro, BrazilReviewed by:
David Marjanović, Museum für Naturkunde, GermanyXiaoke Qiang, Chinese Academy of Sciences, China
Copyright © 2019 Sun and Deng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tao Deng, ZGVuZ3Rhb0BpdnBwLmFjLmNu