 Ivett Pipoly1*
Ivett Pipoly1* Krisztián Szabó2
Krisztián Szabó2 Veronika Bókony3Bálint Preiszner4
Veronika Bókony3Bálint Preiszner4 Gábor Seress5
Gábor Seress5 Erno Vincze1
Erno Vincze1 Julia Schroeder6András Liker1,5
Julia Schroeder6András Liker1,5- 1MTA-PE Evolutionary Ecology Research Group, University of Pannonia, Veszprém, Hungary
- 2Conservation Genetics Research Group, Department of Ecology, University of Veterinary Medicine, Budapest, Hungary
- 3Lendület Evolutionary Ecology Research Group, Plant Protection Institute, Centre for Agricultural Research, Hungarian Academy of Sciences, Budapest, Hungary
- 4Fish Ecology Research Group, MTA ÖK Balaton Limnological Institute, Tihany, Hungary
- 5Department of Limnology, University of Pannonia, Veszprém, Hungary
- 6Department of Life Sciences, Imperial College, London, United Kingdom
Urbanization increasingly changes the ecological conditions for wild animal populations, influencing their demography, reproduction, and behavior. While studies on the ecological consequences of urbanization frequently document a reduced number and poorer body condition of offspring in urban than in non-urban bird populations, consequences for other components of reproduction are rarely investigated. Mating with partners outside the social pair-bond is widespread in birds, and although theory predicts that the occurrence of extra-pair fertilizations (EPF) may be sensitive to the altered ecological conditions of cities, the effect of urbanization on EPF is poorly known. Here we used data from two urban and two forest populations collected over 3 years to test whether the frequency of extra-pair offspring (EPO) in great tit broods differed between the habitats. We found that significantly more broods contained EPO in urban habitats (48.9%) than in forests (24.4%). In broods with EPO, the number and proportion of EPO was similar in urban and forest broods. These results suggest that females that live in urban habitats are more likely to engage in EPF than those living in forests. Urban environments may either provide more spatiotemporal opportunities to EPF because of higher breeding density, and/or enhance motivation for EPF to increase fertility in polluted environments. In addition, females with higher propensity to engage in EPF may more likely settle in urban habitats.
Introduction
Urban animals often differ in their behavior, life history, demographics, and fitness from conspecifics living in more natural habitats (Gil and Brumm, 2013; Rodewald and Gehrt, 2014; Seress and Liker, 2015). In birds, many species successfully colonized urban areas worldwide, and urban individuals have to cope with several anthropogenic environmental changes such as noise, light, and chemical pollution (Seress and Liker, 2015), habitat fragmentation (Crooks et al., 2004), and ecological challenges such as higher population densities (Moller et al., 2012) and lower availability of natural food (Seress et al., 2018). These differences between urban and natural habitats may alter the costs and benefits of birds' reproductive decisions thereby affecting their behavior. In line with this, urban birds typically show altered reproductive biology including advanced laying dates, reduced brood sizes, higher nest-failure rates, and smaller nestlings (Bailly et al., 2016; Seress et al., 2018) compared to their non-urban conspecifics.
A widespread reproductive behavior among pair-bonding species is the pursuit of extra-pair fertilizations (EPF), which are found in ~90% of socially monogamous bird species (Griffith et al., 2002). EPF can increase the number of offspring for males, and it may grant genetic benefits to females by improving fertilization success and/or offspring quality (Griffith et al., 2002; Charmantier et al., 2004). These adaptive functions of EPF have been supported by empirical data, although not unequivocally (Hsu, 2014; Arct et al., 2015). The frequency of EPF can also be influenced by the spatiotemporal distribution of mating opportunities (Schlicht et al., 2014; García-Navas et al., 2015) and physical environmental factors such as night lighting and anthropogenic noise (Kempenaers et al., 2010; Halfwerk et al., 2011). Many of these potential impacts on extra-pair mating behavior may be affected by the altered ecological conditions of urban habitats. However, very few studies have compared EPF between birds breeding in urban and natural habitats (Moore et al., 2012; Rodriguez-Martínez et al., 2014; Bonderud et al., 2018), and it remains unclear whether and how extra-pair behavior varies with habitat urbanization.
The aim of our study was to test whether the frequency of extra-pair offspring (EPO; i.e., offspring that are not genetically related to the social partner of the female) within and across broods differs between urban and non-urban great tits (Parus major), a common and successfully urbanized passerine species with relatively high EPF rates (García-Navas et al., 2015). To test this we used a data set from two urban and two forest populations from three consecutive breeding seasons.
Methods
We studied great tits breeding in nestboxes at two urban and two forest sites in Hungary from 2012 to 2014. We recorded the number of eggs and nestlings in the nestboxes every 3–4 days from March to the end of June. We captured parent birds and took their nestlings before fledging to collect blood samples for genotyping. In 2013 and 2014 we also collected unhatched eggs and tissue samples from nestlings that were found dead before blood sampling. For further details, see Supplementary Information: Field methods.
We selected 86 first annual broods of marked parents and conducted multi-locus genotyping on the whole families by amplifying 5 microsatellite loci with tri- and tetra-nucleotide repeats (Table S1) using multiplex PCR reactions. Altogether 159 parents (80 males and 79 females) and 851 offspring were genotyped. In a subset of samples (n = 23 individuals out of total 1010) with ambiguous results based on the 5 loci, we used 3 additional loci (Table S1). Fluorescent PCR products were scanned by capillary electrophoresis, and alleles were identified and scored by two independent researchers who were blind to the identity of birds. Our marker set proved reliable and efficient for identifying within-pair offspring (WPO) and EPO (see details in Supplementary Information: Genotyping). We identified an offspring as EPO if it mismatched the alleles of the social father on at least two loci but it had no mismatch with the maternal alleles.
We tested the difference in EPO frequencies between urban and forest broods by Generalized Estimation Equations (GEE) models to accommodate the non-normal error distributions and the few non-independent broods, as the majority of broods were independent but a few had one or two parents in common (Zuur et al., 2009). We investigated three response variables: EPO occurrence (EPO present/absent in the brood), the number of EPO per brood, and the proportion of EPO within the brood, the latter calculated as EPO / (EPO + WPO). The model for EPO occurrence contained all 86 broods, while the models for EPO number and EPO proportion contained only those 32 broods where at least one EPO occurred. We chose this subset in the latter models to avoid zero inflation, and to explicitly address the question whether urban and forest parents that engage in EPF differ in their allocation into extra-pair offspring. The predictor variables were study site (4 sites) and year (3 years) in all models. We calculated a linear contrast from each GEE model's estimates to statistically compare the two habitat types (i.e., two urban sites vs. two forest sites). For further details, see Supplementary Information: Statistical analyses.
Results
There were more broods containing EPO in urban than forest habitat (Figure 1; Table 1) in all 3 study years (urban vs. forest 2012: 40.00 vs. 33.33%; 2013: 39.13 vs. 6.67%; 2014: 64.71 vs. 35.29%). When we controlled for non-independence of parents and differences among years in a GEE model (Table S2), EPO occurrence was significantly higher in urban than forest broods [odds ratio calculated from urban-forest linear contrast = 5.77, 95% confidence interval (CI): 1.72–19.4, p = 0.005].
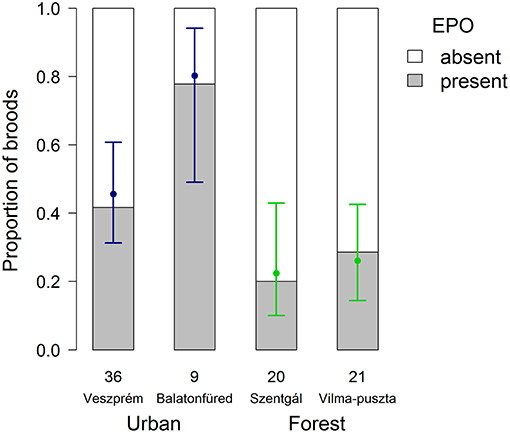
Figure 1. Occurrence of extra-pair offspring (EPO) in great tit broods at urban and forest study sites. Numbers below the bars refer to the number of genotyped broods in each site. Dots and whiskers show means and 87% confidence intervals, respectively, both calculated from the GEE model with study sites and years as predictors. Non-overlapping 87% CIs indicate statistically significant difference (i.e., that a 95% CI of the difference excludes zero).
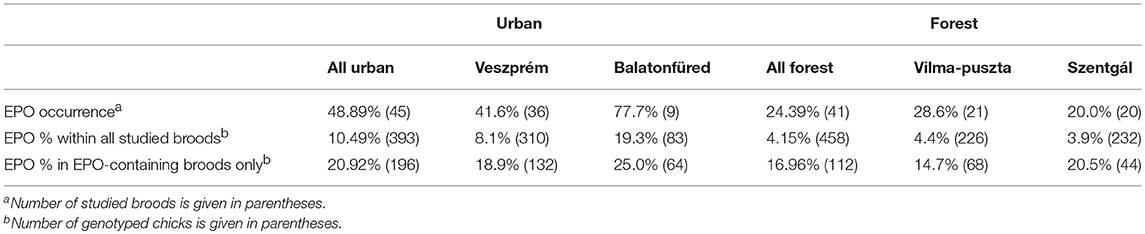
Table 1. Occurrence of extra-pair offspring (EPO), i.e., proportion of studied broods that contained at least one EPO, and the percentage of EPO among all nestlings in the two urban and two forest study sites.
The number and proportion of EPO was slightly higher in urban broods than in forests when we considered all broods (Table 1), but when we considered only the broods with at least one EPO there was no difference between urban and forest habitats in the number of EPO per brood (Table 1; Figure S1; Table S2, proportional difference between urban and forest broods from linear contrast = 1.175, 95% CI: 0.667–2.067, p = 0.58) and the proportion of EPO per brood (Table 1; Figure S2; Table S2, odds ratio from urban-forest linear contrast = 1.65, 95% CI: 0.793–3.43, p = 0.18). The results for all three variables of interest remained the same when we omitted data from 2012 when we did not have data for one urban site and did not collect samples from dead chicks and unhatched eggs (Supplementary Material, Table S3).
Discussion
We found that urban broods of great tits contained at least one EPO significantly more often than forest broods, and this pattern was consistent across all 3 years. This result corroborates findings on other species: EPF tended to be higher in more urbanized areas in Canada geese (Branta canadensis) (Moore et al., 2012) and an unusually high rate of EPF was registered in urban Cooper's hawks (Accipiter cooperii) (Rosenfield et al., 2015). However, there was no habitat difference in EPF rate of mountain chickadees (Poecile gambeli) (Bonderud et al., 2018). On the other hand, in broods where EPO occurred we did not find difference in the number and proportion of EPO between urban and forest habitats. Thus, our results suggest that urbanization is associated with increased occurrence of EPF in great tits, although those females that engage in EPF produce similar numbers of EPO in both habitat types.
Several factors might explain the higher occurrence of EPO in urban populations. First, urban sites may offer more opportunities in space and time, for example through higher breeding density, as urban birds on average have 30% higher breeding density than rural populations (Moller et al., 2012), and higher breeding density can increase EPF frequency (Charmantier and Perret, 2004; Mayer and Pasinelli, 2013). Moreover, urban individuals often have a prolonged diurnal activity due to artificial lighting at night (Dominoni et al., 2013), offering more time for females to search for extra-pair males during early dawn (Double and Cockburn, 2000; Kempenaers et al., 2010; Halfwerk et al., 2011). Furthermore, night lighting enables males to start singing earlier and such males are more successful in siring EPO (Silva et al., 2015).
Further, food availability and its seasonal distribution may also contribute to the differences in extra-pair mating behavior between urban and forest populations. Caterpillars, the main food of great tits during the breeding season, are scarce in cities (Biard et al., 2017; Seress et al., 2018), so urban males might have to spend more time foraging at the expense of guarding their females during their fertile period, increasing opportunities for EPF in cities. However, previous experimental studies found that food scarcity can either increase (Hoi-leitner et al., 1999) or decrease EPF rates in various species (Václav et al., 2003; Kaiser et al., 2015). Furthermore, urban habitats lack the massive spring peak of caterpillar abundance that is typical in forests (Seress et al., 2018), which might lead to less synchronized breeding in urban populations, further facilitating EPF (Van Dongen and Mulder, 2009; García-Navas et al., 2015), although an opposite effect of breeding asynchrony is also possible (Stutchbury and Morton, 1995; Neudorf, 2004).
Also, birds might gain more benefits from EPF in urban habitats than in forests. Urban birds can suffer from a higher risk of inbreeding (Vangestel et al., 2011; Rodriguez-Martínez et al., 2014) and reduced reproductive success per breeding attempt (Bailly et al., 2016; Biard et al., 2017; Seress et al., 2018), so any benefit gained from fertility insurance and good genes might have more value in urban areas (Reding, 2014). Furthermore, urban females may be more motivated to pursue extra-pair partners because they might be more likely to perceive their social mate's quality as low. Individual quality and/or signals like song and plumage ornaments in birds are often relatively poorly developed in urban habitats, including the great tits' yellow plumage coloration (Halfwerk et al., 2011; Biard et al., 2017), black breast stripe (Senar et al., 2014), and song characteristics (Slabbekoorn and Peet, 2003). Urban great tit males that sing higher-frequency songs to overcome the noise-induced communication breakdown are cuckolded more often (Halfwerk et al., 2011), although another study found no difference in EPF in house sparrows (Passer domesticus) between noisy and quiet breeding sites (Schroeder et al., 2012).
Alternatively, the relationship between urbanization and EPO occurrence might be non-causal. For example, individuals with different behavioral types can differ in their habitat choice (Holtmann et al., 2017; Sprau and Dingemanse, 2017) and the propensity to engage in EPF can vary between females as part of their personality (Forstmeier, 2007; Forstmeier et al., 2014; Wolak et al., 2018). It is thus possible that a tendency for promiscuity is associated with the behavioral traits that facilitate settlement in cities, such as innovative problem solving and exploratory behavior (Bókony et al., 2017).
Taken together, there are many conceivable mechanisms by which habitat urbanization may influence the frequency of EPF. Understanding how these mechanisms shape avian reproductive behaviors, and why their effects differ between species, will further expand our knowledge on urban behavioral ecology and sexual selection. Furthermore, because EPF can decrease inbreeding and increase genetic diversity, uncovering the genetic mating systems of urban animals and identifying the main factors of urban cuckoldry may aid the conservation of fragmented populations in our urbanizing world.
Data Availability
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
This study was carried out in accordance with the recommendations for animal care outlined by ASAB/ABS (www.asab.org) and Hungarian laws, licensed by the Government Office of Veszprém County, Nature Conservation Division (former Middle Transdanubian Inspectorate for Environmental Protection, Natural Protection and Water Management; permission number: 31559/2011).
Author Contributions
IP, VB, KS, and AL designed the study. IP, VB, BP, GS, EV, and AL collected data in the field. IP and KS did the molecular work and read the genotypes. KS made the parentage analyses. IP, VB, and AL did the statistical analyses. All authors wrote the manuscript.
Funding
The project was supported by the European Union, with the co-funding of the European Social Fund (TÁMOP-4.2.2.A-11/1/KONV-2012-0064) and by the Hungarian Scientific Research Fund (NKFIH K84132, K112838). IP was supported by the ÚNKP-17-3 New National Excellence Program of the Ministry of Human Capacities, Hungary. VB was supported by the János Bolyai Scholarship of the Hungarian Academy of Sciences. GS was supported by an NKFIH postdoctoral grant (PD 120998) during the preparation of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank T. Hammer and S. Papp for helping with field work. We also thank the reviewers for useful suggestions to improve our paper.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00229/full#supplementary-material
References
Arct, A., Drobniak, S. M., and Cichon, M. (2015). Genetic similarity between mates predicts extrapair paternity—a meta-analysis of bird studies. Behav. Ecol. 26, 959–968. doi: 10.1093/beheco/arv004
Bailly, J., Scheifler, R., Berthe, S., Clément-Demange, V.-A., Leblond, M., Pasteur, B., et al. (2016). From eggs to fledging: negative impact of urban habitat on reproduction in two tit species. J. Ornithol. 157, 377–392. doi: 10.1007/s10336-015-1293-3
Biard, C., Brischoux, F., Meillère, A., Michaud, B., and Nivière, M. (2017). Growing in cities: an urban penalty for wild birds? A study of phenotypic differences between urban and rural great tit chicks (Parus major). Front. Ecol. Evol. 5:79. doi: 10.3389/fevo.2017.00079
Bókony, V., Pipoly, I., Szabó, K., Preiszner, B., Vincze, E., Papp, S., et al. (2017). Innovative females are more promiscuous in great tits (Parus major). Behav. Ecol. 28, 579–588. doi: 10.1093/beheco/arx001
Bonderud, E. S., Otter, K. A., Burg, T. M., Marini, K. L. D., and Reudink, M. W. (2018). Patterns of extra-pair paternity in mountain chickadees. Ethology 124, 378–386. doi: 10.1111/eth.12747
Charmantier, A., Blondel, J., Perret, P., and Lambrechts, M. M. (2004). Do extra-pair paternities provide genetic benefits for female blue tits Parus caeruleus? J. Avian Biol. 35, 524–535. doi: 10.1111/j.0908-8857.2004.03296.x
Charmantier, A., and Perret, P. (2004). Manipulation of nest-box density affects extra-pair paternity in a population of blue tits (Parus caeruleus). Behav. Ecol. Sociobiol. 56, 360–365. doi: 10.1007/s00265-004-0794-5
Crooks, K. R., Suarez, A. V., and Bolger, D. T. (2004). Avian assemblages along a gradient of urbanization in a highly fragmented landscape. Biol. Conserv. 115, 451–462. doi: 10.1016/S0006-3207(03)00162-9
Dominoni, D. M., Helm, B., Lehmann, M., Dowse, H. B., and Partecke, J. (2013). Clocks for the city : circadian differences between forest and city songbirds. Proc. R. Soc. B 280, 1–7. doi: 10.1098/rspb.2013.0593
Double, M., and Cockburn, A. (2000). Pre-dawn infidelity: females control extra-pair mating in superb fairy-wrens. Proc. R. Soc. B 267, 465–470. doi: 10.1098/rspb.2000.1023
Forstmeier, W. (2007). Do individual females differ intrinsically in their propensity to engage in extra-pair copulations? PLoS ONE 2:e952. doi: 10.1371/journal.pone.0000952
Forstmeier, W., Nakagawa, S., Griffith, S. C., and Kempenaers, B. (2014). Female extra-pair mating: adaptation or genetic constraint? Trends Ecol. Evol. 29, 456–464. doi: 10.1016/j.tree.2014.05.005
García-Navas, V., Ferrer, E. S., Cáliz-Campal, C., Bueno-Enciso, J., Barrientos, R., Sanz, J. J., et al. (2015). Spatiotemporal and genetic contingency of extrapair behaviour in a songbird. Anim. Behav. 106, 157–169. doi: 10.1016/j.anbehav.2015.05.020
Gil, D., and Brumm, H. (2013). Avian Urban Ecology: Behavioural and Physiological Adaptations. Oxford: Oxford University Press. doi: 10.1093/acprof:osobl/9780199661572.001.0001
Griffith, S. C., Owens, I. P. F., and Thuman, K. A. (2002). Extra pair paternity in birds : a review of interspecific variation and adaptive function. Mol. Ecol. 11, 2195–2212. doi: 10.1046/j.1365-294X.2002.01613.x
Halfwerk, W., Bot, S., Buikx, J., van der Velde, M., Komdeur, J., ten Cate, C., et al. (2011). Low-frequency songs lose their potency in noisy urban conditions. Proc. Natl. Acad. Sci. U.S.A. 108, 14549–14554. doi: 10.1073/pnas.1109091108
Hoi-leitner, M., Hoi, H., Romero-Pujante, M., and Valera, F. (1999). Female extra-pair behaviour and environmental quality in the serin (Serinus serinus): a test of the “constrained female hypothesis.” Proc. R. Soc. B 266, 1021–1026. doi: 10.1098/rspb.1999.0738
Holtmann, B., Santos, E. S. A., Lara, C. E., and Nakagawa, S. (2017). Personality-matching habitat choice, rather than behavioural plasticity, is a likely driver of a phenotype – environment covariance. Proc. R. Soc. B 284, 1–9. doi: 10.1098/rspb.2017.0943
Hsu, Y. (2014). The Causes and Consequences of Extra - Pair Mating in House Sparrows (Passer domesticus) (Thesis, Doctor of Philosophy). University of Otago, 176.
Kaiser, S. A., Sillett, T. S., Risk, B. B., and Webster, M. S. (2015). Experimental food supplementation reveals habitat-specific reproductive investment in a migratory bird. Proc. R. Soc. B 282:20142523. doi: 10.1098/rspb.2014.2523
Kempenaers, B., Borgström, P., Loës, P., Schlicht, E., and Valcu, M. (2010). Artificial night lighting affects dawn song, extra-pair siring success, and lay date in songbirds. Curr. Biol. 20, 1735–1739. doi: 10.1016/j.cub.2010.08.028
Mayer, C., and Pasinelli, G. (2013). New support for an old hypothesis: density affects extra-pair paternity. Ecol. Evol. 3, 694–705. doi: 10.1002/ece3.489
Moller, A. P., Diaz, M., Flensted-Jensen, E., Grim, T., Ibánez-Álamo, J. D., Jokimaki, J., et al. (2012). High urban population density of birds reflects their timing of urbanization. Oecologia 170, 867–875. doi: 10.1007/s00442-012-2355-3
Moore, J. A., Kamarainen, A. M., Scribner, K. T., Mykut, C., and Prince, H. H. (2012). The effects of anthropogenic alteration of nesting habitat on rates of extra-pair fertilization and intraspecific brood parasitism in Canada Geese Branta canadensis. Ibis 154, 354–362. doi: 10.1111/j.1474-919X.2011.01203.x
Neudorf, D. L. H. (2004). Extrapair paternity in birds: understanding variation among species. Am. Nat. 121, 302–307. doi: 10.1642/0004-8038(2004)121[0302:EPIBUV]2.0.CO;2
Reding, L. (2014). Increased hatching success as a direct benefit of poliandry in birds. Evolution 69, 264-270. doi: 10.1111/evo.12553
Rodewald, A. D., and Gehrt, S. D. (2014). “Wildlife population dynamics in urban landscapes,” in Urban Wildlife Conservation: Theory and Practice, eds R. A. McCleery, C. E. Moorman, and M. N. Peterson (New York, NY: Springer Science+Business Media), 117–147. doi: 10.1007/978-1-4899-7500-3_8
Rodriguez-Martínez, S., Carrete, M., Roques, S., Rebolo-Ifrán, N., and Tella, J. L. (2014). High urban breeding densities do not disrupt genetic monogamy in a bird species. PLoS ONE 9:e91314. doi: 10.1371/journal.pone.0091314
Rosenfield, R. N., Sonsthagen, S. A., Stout, W. E., and Talbot, S. L. (2015). High frequency of extra-pair paternity in an urban population of Cooper's Hawks. J. Field Ornithol. 86, 144–152. doi: 10.1111/jofo.12097
Schlicht, L., Valcu, M., and Kempenaers, B. (2014). Spatial patterns of extra-pair paternity: beyond paternity gains and losses. J. Anim. Ecol. 84, 518–531. doi: 10.1111/1365-2656.12293
Schroeder, J., Nakagawa, S., Cleasby, I. R., and Burke, T. (2012). Passerine birds breeding under chronic noise experience reduced fitness. PLoS ONE 7:e39200. doi: 10.1371/journal.pone.0039200
Senar, J. C., Conroy, M. J., Quesada, J., and Mateos-gonzalez, F. (2014). Selection based on the size of the black tie of the great tit may be reversed in urban habitats. Ecol. Evol. 4, 2625–2632. doi: 10.1002/ece3.999
Seress, G., Hammer, T., Bókony, V., Vincze, E., Preiszner, B., Pipoly, I., et al. (2018). Impact of urbanization on abundance and phenology of caterpillars and consequences for breeding in an insectivorous bird. Ecol. Appl. 28, 1143–1156. doi: 10.1002/eap.1730
Seress, G., and Liker, A. (2015). Habitat urbanization and its effects on birds. Acta Zool. Acad. Sci. Hung. 61, 373–408. doi: 10.17109/AZH.61.4.373.2015
Silva, A., Valcu, M., and Kempenaers, B. (2015). Light pollution alters the phenology of dawn and dusk singing in common European songbirds. Philos. Trans. R. Soc. Lond. B Biol. Sci. 370, 1–9. doi: 10.1098/rstb.2014.0126
Slabbekoorn, H., and Peet, M. (2003). Birds sing at a higher pitch in urban noise. Nature 424, 267. doi: 10.1038/424267a
Sprau, P., and Dingemanse, N. J. (2017). An approach to distinguish between plasticity and non-random distributions of behavioral types along urban gradients in a wild passerine bird. Front. Ecol. Evol. 5:92. doi: 10.3389/fevo.2017.00092
Stutchbury, B. J., and Morton, E. S. (1995). The effect of breeding synchrony on extra-pair mating systems in songbirds. Behaviour 132, 675–690. doi: 10.1163/156853995X00081
Václav, R., Hoi, H., and Blomqvist, D. (2003). Food supplementation affects extrapair paternity in house sparrows (Passer domesticus). Behav. Ecol. 14, 730–735. doi: 10.1093/beheco/arg059
Van Dongen, W. F. D., and Mulder, R. A. (2009). Multiple ornamentation, female breeding synchrony, and extra-pair mating success of golden whistlers (Pachycephala pectoralis). J. Ornithol. 150, 607–620. doi: 10.1007/s10336-009-0371-9
Vangestel, C., Mergeay, J., Dawson, D. A., Vandomme, V., and Lens, L. (2011). Spatial heterogeneity in genetic relatedness among house sparrows along an urban-rural gradient as revealed by individual-based analysis. Mol. Ecol. 20, 4643–4653. doi: 10.1111/j.1365-294X.2011.05316.x
Wolak, M. E., Arcese, P., Keller, L. F., Nietlisbach, P., and Reid, J. M. (2018). Sex-specific additive genetic variances and correlations for fitness in a song sparrow (Melospiza melodia) population subject to natural immigration and inbreeding. Evolution 72, 2057–2075. doi: 10.1111/evo.13575
Keywords: extra-pair paternity, promiscuity, cuckoldry, urbanization, reproductive behavior
Citation: Pipoly I, Szabó K, Bókony V, Preiszner B, Seress G, Vincze E, Schroeder J and Liker A (2019) Higher Frequency of Extra-Pair Offspring in Urban Than Forest Broods of Great Tits (Parus major). Front. Ecol. Evol. 7:229. doi: 10.3389/fevo.2019.00229
Received: 11 March 2019; Accepted: 03 June 2019;
Published: 20 June 2019.
Edited by:
François Criscuolo, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Sylvie Massemin, UMR7178 Institut Pluridisciplinaire Hubert Curien (IPHC), FranceF. Stephen Dobson, Auburn University, United States
Copyright © 2019 Pipoly, Szabó, Bókony, Preiszner, Seress, Vincze, Schroeder and Liker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ivett Pipoly, cGlwb2x5Lml2ZXR0JiN4MDAwNDA7bWsudW5pLXBhbm5vbi5odQ==