 Raquel Muñoz-Gallego
Raquel Muñoz-Gallego José M. Fedriani
José M. Fedriani Anna Traveset
Anna Traveset- 1Department of Physical, Chemical and Natural Systems, University of Pablo de Olavide, Seville, Spain
- 2Department of Conservation Biology, Doñana Biological Station (EBD-CSIC), Seville, Spain
- 3Global Change Research Group, Mediterranean Institute of Advanced Studies (IMEDEA- CSIC), Esporles, Spain
- 4Desertification Research Centre CIDE, CSIC-UVEG-GV, Moncada, Spain
- 5Centre for Applied Ecology “Prof. Baeta Neves”, InBio, Institute Superior of Agronomy, University of Lisbon, Lisbon, Portugal
Megafauna extinctions often lead to the disruption of plant-animal interactions, such as the seed-disperser mutualisms, which might entail severe consequences for plant populations and entire communities. Interestingly, the contemporary persistence of anachronistic plant species might be possible thanks to surrogate dispersers or seed dispersal “rescuers”. We know very little on how these relevant functional replacements are contributing to the performance of present-day plant-frugivore networks. The dwarf palm Chamaerops humilis L. is a Mediterranean endemism with fleshy fruits and typically dispersed by mammals. Despite its ecological importance and wide distribution in some of the Mediterranean islands, no information exists about its seed dispersal on these depauperated-fauna systems. In this study, we aim at identifying and quantifying the relative importance of introduced frugivores on the island of Mallorca (Balearic Islands), where no native terrestrial mammals exist. Specifically, we assess for the first time the seed dispersal effectiveness (SDE) for C. humilis on islands; we evaluate the quantitative component by fecal and regurgitation sampling surveys, and the qualitative component by means of seed germination experiments and seedling growth measures. Introduced goats (Capra hircus L.) and pine martens (Martes martes L.) were the local mammal fruit consumers of C. humilis identified in our study sites. Results suggest that goats are much more important quantitatively than pine martens, due to the high number of fruits handled in each foraging bout and their extremely high abundance on the island. However, pine marten-ingested seeds showed the highest final seedling emergence success and seedling growth, thus its qualitative contribution on C. humilis seed dispersal is higher than that of goats. Overall, SDE was almost 9-fold higher for goats than for pine martens. We conclude that these two non-native mammal species are effective seed dispersers of C. humilis in this and probably other Mediterranean islands, where humans led to the extinction of its native seed dispersers, as it was probably the case of the goat-like Myotragus balearicus in the Balearic Islands.
Introduction
Numerous extinctions of large terrestrial vertebrates driven by human activity have taken place during the Late Pleistocene and Early Holocene (Ceballos et al., 2015; Faurby and Svenning, 2015). This has occurred in most continents, and well-studied cases are the extinctions of megafauna species in the American continent, such as saber-toothed cats (Smilodon spp.), mammoths (Mammuthus spp.), and giant ground sloths (Megalonyx jeffersonii) (Janzen and Martin, 1982; Sodhi et al., 2012). On islands worldwide, the rate of extinction of terrestrial mammals after the arrival of humans was c. 30%, being fateful for the orders Proboscidea, Xenarthra and Bibymalagasia and quite strong for Artiodactyla, Carnivora, and the rodent infraorder Hystricognathi (Alcover et al., 1998). These events not only led to the vanishing of many populations and species, but also to the disruption and reshaping of ecological interactions which likely altered the structure and function of entire communities and ecosystems. In fact, severe impacts of large vertebrate extinctions on the ecosystem physical and trophic structure, plant community composition, and diversity, ecosystem biogeochemistry, and regional and global climate have been described (Dirzo et al., 2014; Bello et al., 2015; Ripple et al., 2015; Malhi et al., 2016; Smith et al., 2016). In addition, the loss of megafauna can affect ecological interactions in three ways: leading to co-extinctions, adaptative shifting, and evolutionary anachronisms (Galetti et al., 2018). This last concept, described for the first time several decades ago by Janzen and Martin (1982), is defined in a context of frugivory as “extant interactions between animal frugivores and plants involving traits that show striking unfit patterns to an extant fauna” (Guimarães et al., 2008, p. 2). Typical examples are fruits with huge seeds which are dispersed inefficiently by the contemporary frugivore assemblages, like avocado (Persea americana) (Cook, 1982), osage orange (Maclura pomifera) (Janzen and Martin, 1982), or Florida torreya (Torreya taxifolia) (Barlow, 2001). Anachronisms can also show up through antagonisms, such as the presence of spines or other plant defenses against large herbivores that no longer exist (e.g., Bond et al., 2004). These unfit patterns could be explained if we consider all the extinct megafauna which would have acted as legitimate dispersers or herbivores in the past, exerting selective pressures on plant traits (Barlow, 2000; Guimarães et al., 2008).
Seed dispersal is a key ecological process in the life cycle of plants because it allows propagules to move away from the maternal environment, providing a higher propagule survival probability, maintaining the genetic flow, and enabling the colonization of vacant habitats (Levey et al., 2002; Medel et al., 2009; Schupp et al., 2010). Therefore, the disruption of this relevant ecological service due to the extinction of effective seed dispersers can result in serious consequences for plant populations: modification of range and spatial distribution (e.g., Meehan et al., 2002; Rotllàn-Puig and Traveset, 2016), decrease in the total number of seed successfully dispersed, germinated and established (e.g., Galetti et al., 2006; Wotton and Kelly, 2011; Traveset et al., 2012), alteration of the seed shadow (e.g., Beaune et al., 2013; Bueno et al., 2013), restriction of the gene flow and genetic structure increase (e.g., Collevatti et al., 2003; Caughlin et al., 2015), interruption of long-distance dispersal (e.g., Pérez-Méndez et al., 2016; Pires et al., 2018), and shift of phenotypic selection on seed size (e.g., Galetti et al., 2013). This often ends in a collapse in the life cycle and species decline (Valiente-Banuet et al., 2015; Rumeu et al., 2017; Galetti et al., 2018). On islands, where communities are depauperate and plants may be very specialized in their dispersers (González-Castro et al., 2012), the negative outcomes from such disruptions are even greater (Traveset and Richardson, 2014). The disappearance of frugivorous forest birds (e.g., moas, piopio Turnagra capensis, huia Heterolocha acutirostris) as important dispersers of many woody plants on New Zealand (Clout and Hay, 1989; Wood et al., 2008), or the reduction of flying foxes on tropical Pacific islands (McConkey and Drake, 2006) are some examples of disrupted dispersal service on islands, although the list is long and increases (e.g., Rogers et al., 2017; Rumeu et al., 2017). In the Mediterranean basin, few studies have been carried out on the disruption of plant-disperser mutualisms (e.g., Traveset, 2002; Traveset et al., 2012). Interestingly, the persistence of large-fleshy-fruited species has often been possible thanks to extant small-sized frugivores mammals such as rodents (Jansen et al., 2012), and also to introduced carnivores or livestock (Janzen and Martin, 1982), which may act as “surrogate Pleistocene dispersal agents” (sensu Janzen, 1982). Other examples have been documented from other systems. For instance, in the Balearic Islands, introduced pine martens (Martes martes) replace, at least partially, the native lizards which presumably used to be the only legitimate dispersers of the native shrub Cneorum tricoccon until recently (Traveset, 1995; Celedón-Neghme et al., 2013). Therefore, contemporary frugivore assemblages, even though in a suboptimal way (Guimarães et al., 2008), can act as “rescuers” of the seed dispersal service of anachronistic species, mitigating the effects of previous animal extinctions, and avoiding the vanishing of ecological interactions (Zamora, 2000; Valiente-Banuet et al., 2015; Rumeu et al., 2017; Cares et al., 2018).
On Mallorca island (Balearic archipelago), the native vertebrate fauna of the Upper Pleistocene and the Holocene was composed only of three endemic species: the cave goat Myotragus balearicus (Artiodactyla: Bovidae; 25.19–33.34 kg, Palombo et al., 2008), the giant dormouse Hypnomys morpheus (Rodentia: Gliridae; 0.23 kg, Moncunill-Solé et al., 2014) and the Balearic shrew Nesiotites hidalgo (Soricomorpha: Soricidae; 0.023–0.031 kg, Bover et al., 2008; Moncunill-Solé et al., 2016). The cave goat was thus the only medium-sized herbivorous mammal present on the islands (specifically, on Mallorca, Menorca and Cabrera). Its extinction took place during the Holocene, around 3000–2000 BC, and has been associated to the first arrival of humans to these islands (Bover and Alcover, 2008; Bover et al., 2016). Nowadays, the only non-flying mammals present in the Balearics have all been introduced by man, including goats, sheep, cows, pigs, dogs, as well as rodents (Bover and Alcover, 2008; Valenzuela and Alcover, 2013a). The wild carnivore the weasel (Mustela nivalis) appears to have been imported by Talaiotic people (prior to 123 BC) (Valenzuela and Alcover, 2013b). Subsequently, during the Roman period (123 BC-534 AD), the domestic cat (Felis catus) and the pine marten (M. martes) were introduced (Valenzuela and Alcover, 2015), and the common genet (Genetta genetta) was also probably introduced previously to the Muslims arrival (656–773 AD; Delibes et al., 2017b). The raccoon (Procyon lotor) and the coati (Nasua nasua) have been established very recently (in 2006 and 2004, respectively) on the island (Mayol et al., 2009; Pinya et al., 2009). However, very little is known about the ecological role of these introduced vertebrate species on the dynamics and composition of island communities.
In the present study, we aim at identifying and quantifying the relative importance of introduced seed dispersers for the dwarf palm Chamaerops humilis L. on Mallorca. This endemic palm is known to be mammal-dispersed elsewhere (Fedriani and Delibes, 2011) and we thus expect that non-native mammals play the role of dispersers in this island. In the few areas of continental Europe where its seed dispersal has been studied (Table 1), carnivorous mammals such as the European badger (Meles meles) and the red fox (Vulpes vulpes) have been described as its legitimate dispersers, and rabbits (Oryctolagus cuniculus) and some ungulates, like the wild boar (Sus scrofa), deer (Cervidae) and the domestic goat (C. hircus), act as occasional and suboptimal dispersal agents (Fedriani and Delibes, 2008, 2011; Delibes et al., 2017a). On islands, for which information is less available, only a few species have been described as consumers of the palm fruits (see Table 1). In Mallorca, its seeds have only been documented from G. genetta feces (Alcover, 1984; Clevenger, 1995), and no study has been carried out on the seed dispersal of this plant despite its high ecological, cultural and economic value (Guzmán et al., 2017).
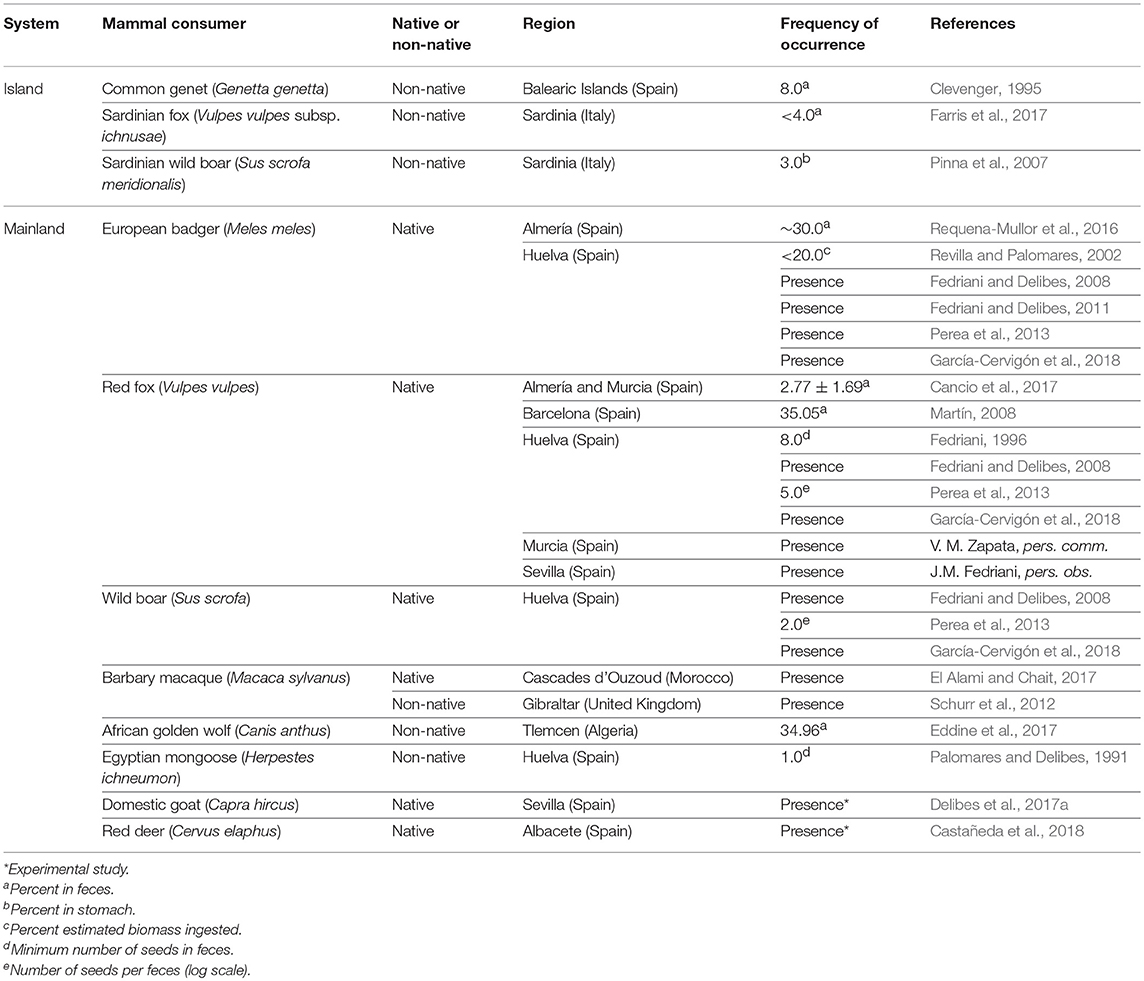
Table 1. Mammal species that consume C. humilis fruits described in the Mediterranean basin so far, indicating the system (island or mainland), the region, whether they are native or not, the frequency of occurrence of dwarf palm fruits in their diet, and the reference of the study.
The specific questions addressed in this study are the following: (1) Who are currently the main vertebrate fruit consumers of C. humilis on Mallorca? (2) How quantitatively and qualitatively effective are such fruit consumers as seed dispersers? To estimate the quantitative component of the seed dispersal effectiveness (SDE) of potential seed dispersers, we searched for mammal droppings and regurgitations, throughout linear transects in areas where this plant is abundant. On the other hand, the qualitative component of SDE for each seed disperser was assessed by means of a seed germination experiment using C. humilis seeds extracted from mammal samples as well as control (i.e., non-ingested) seeds. From our results, we discuss on the functional role of non-native mammal species as seed dispersers of this ancient Mediterranean palm on islands.
Materials and Methods
Study Area
The study was carried out in the Northeast of the Mallorca island, specifically in the North of Serra de Tramuntana (municipality of Pollença) and in the North of Serra de Llevant (municipality of Artá). Four study sites, called “Cala Bóquer”, “Cap de Formentor”, “Ermita de Betlem” and “Es Caló” (Figure 1), were chosen. The predominant forest species are Aleppo pine (Pinus halepensis) and oak (Quercus ilex). Several shrub species, such as C. humilis, Olea europaea var. sylvestris, Pistacia lentiscus, Erica multiflora, Cistus monspeliensis, Ampelodesmus mauritanicus, and Calicotome spinosa are abundant over this territory. The climate of the Balearic archipelago is typically Mediterranean, characterized by two rainy seasons, a hot dry summer and a soft winter. Monthly rainfall on Mallorca varied between 0 and 107.2 mm during 2017, with the most rain falling in January and September, and extreme drought in May. Annual average temperature was 18.2°C (data from Red de Estaciones Meteorológicas de Baleares1).
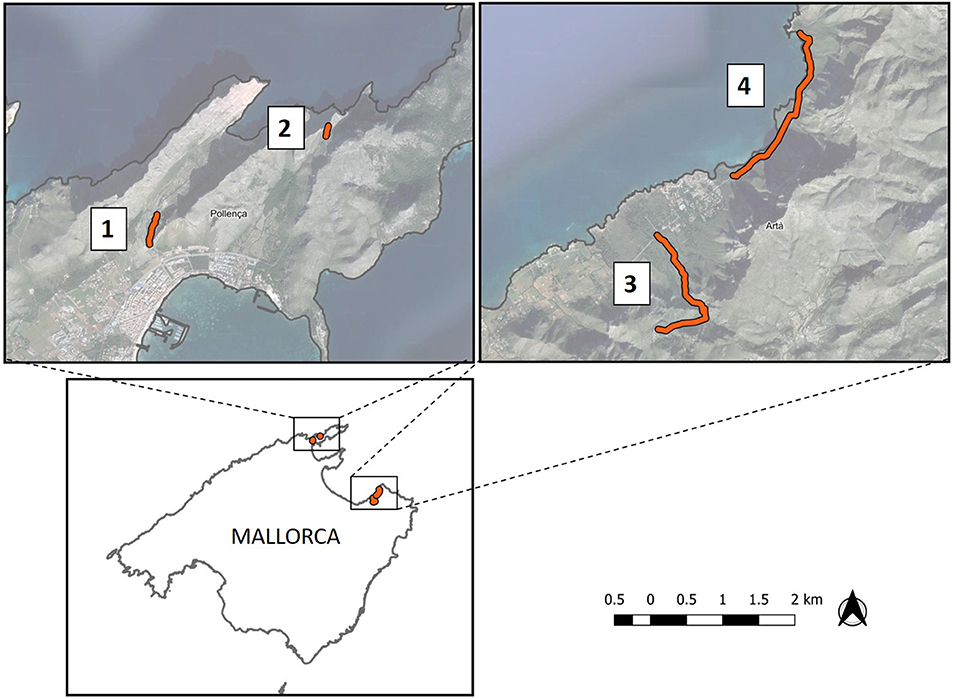
Figure 1. Study area located in the Northeast of the Mallorca island. Orange lines are the four linear transects used for the fecal and regurgitation sampling surveys: (1) Cala Bóquer, (2) Cap de Formentor, (3) Ermita de Betlem, (4) Es Caló.
Study Species
The endemic Mediterranean dwarf palm Chamaerops humilis L. (Arecaceae) is a representative species of the Pre-Pliocene paleo-tropical ancestral lineages (Thompson, 2005). It is very likely that this ancient palm was in the Balearic Islands long before the arrival of the first settlers since its presence in the eastern coast of the Iberian Peninsula (Arroyo et al., 2004; Pérez-Obiol et al., 2010) and in other Mediterranean islands (Sardinia; Biondi and Filigheddu, 1990) is very old, previous to the Mediterranean Sea desiccation. Nevertheless, this remains uncertain so far due to the lack of any pre-anthropic evidence in both pollen and fossil records (Carrión, 2012), which is not surprising because of the poor preservation of this species and the little amount of pollen that scatters (F. Burjachs, pers. comm.) due to, probably, its insect-pollinated syndrome (Carrión, 2002). In Mallorca, this dioecious palm is relatively abundant currently, especially in Serra de Tramuntana and in the North of Serra de Llevant (Garcia and Morey, 1989; Bioatles—Govern de les Illes Balears2). It flowers during spring (March–May) and the fruits ripen in autumn (September–November). Its fleshy fruits are poly-drupes (Herrera, 1989). Unripe fruits are bright green, turning to dull yellow to brown when ripe. The seed (usually 0.6– 0.8 g, 16.3 × 11.0 mm average dimensions, Delibes et al., 2017a) comprises a small cylindrical embryo (2 mg), which is surrounded by several layers, from inner to outer: (1) a nutritious endosperm, (2) a wide woody layer or endocarp, (3) a fleshy and fibrous mesocarp (the pulp, that smells strongly of rancid butter when ripe), and (4) the thin outer layer or exocarp (González-Benito et al., 2006; Hasnaoui et al., 2009). The medium-sized seed is usually large enough to preclude their consumption by birds, so this plant species belongs to the known “mammal-dispersal syndrome”. Occasionally, though, birds may remove some fruits and thus transport seeds to different distances; specifically, one songbird (Turdus merula) was once observed picking one fruit and leaving with it in its bill (R. Muñoz-Gallego, pers. obs.). The dwarf palm seeds are commonly predated by beetles. At least, two non-native scolytine species have been described to bore into C. humilis seeds in the continent, Coccotrypes dactyliperda and Dactylotrypes longicollis, taking place as both pre- and post-dispersal events. Thus, these beetles bore the pulp and endocarp of fruits still attached to the mother plant, dropped from inflorescences, and also of dispersed (and depulped) seeds (Rodríguez et al., 2014; this study). Fruits are attached to infructescences (or ramets) of up to 30 cm long (20–40 fruits per ramet, Fedriani and Delibes, 2011) and located at 10–30 cm from the ground level. Germination is hypogeal and remote, with most seedlings emerging during spring time. Moreover, it has a well-known ability to thrive on poor nutrient soils (Herrera, 1989).
Seed Dispersal Effectiveness: Quantitative and Qualitative Components
Seed dispersal effectiveness (SDE) is defined as the “number of new adult plants produced by the dispersal activities of a disperser” (Schupp, 1993, p. 16). SDE can be quantified as the number of seeds dispersed by a dispersal agent (quantitative effectiveness) multiplied by the probability that a dispersed seed produces a new adult (qualitative effectiveness): SDE = Quantity · Quality. In our study, the quantitative component of SDE was obtained by recording the number of mammal samples that contained C. humilis seeds multiplied by the mean number of seeds per sample (Figure 2). This metric was then corrected by the distance (km) of each transect (e.g., Suárez-Esteban et al., 2013). On the other hand, the qualitative component (i.e., quality of treatment in the mouth and gut) was assessed by recording seedling emergence and seedling growth. The latter was determined by measuring both seedling length and dry weight (Figure 2). Finally, the SDE landscape (sensu Schupp et al., 2010), a visual representation of effectiveness and the relative contribution of each disperser, was estimated by multiplying the quantity component (number of dispersed seeds per distance sampled) by the quality component (emergence success). The landscape was plotted with the code provided by Jordano (2014) with Rstudio v.1.1.383 (R Core Team, 2017).
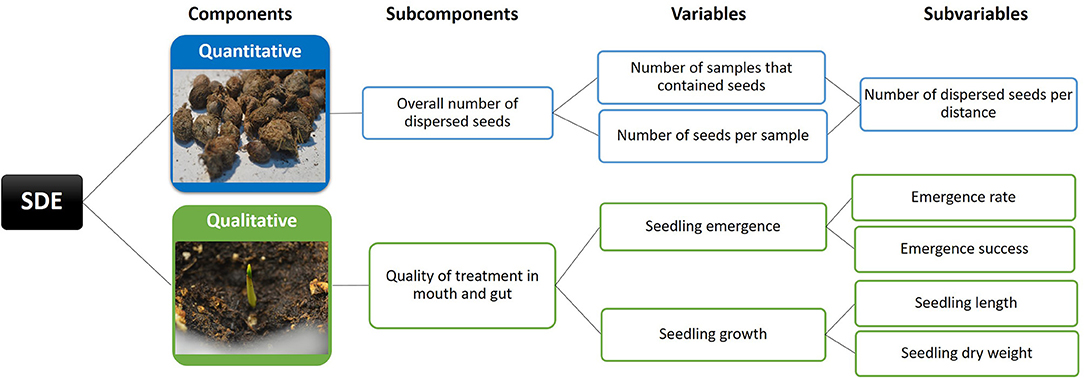
Figure 2. Summary of methods used to assess each of the SDE (Seed Dispersal Effectiveness) components: quantitative and qualitative (sensu Schupp et al., 2010).
Fecal and Regurgitation Sampling Surveys
In November of 2017, we searched for fresh mammal droppings and regurgitations belonging to the second half of the 2017 fruiting season (determined by their non-dry appeareance), along forest tracks and adjacent forest and garrigues. The four transects surveyed are shown in Figure 1: “Cala Bóquer” with 0.44 km in length (start point: 39° 54′ 53.55″ N, 3° 4′ 55.60″ E), “Cap de Formentor” with 0.15 km (start point: 39° 55′ 46.54″ N, 3° 6′ 39.93″ E), “Ermita de Betlem” with 2.25 km (start point: 39° 44′ 56.80″ N, 3° 18′ 41.23″ E), and “Es Caló” with 2.6 km (start point: 39° 45′ 23.60″ N, 3° 19′ 25.80″ E). The first two are located in Serra de Tramuntana and the last two in Serra de Llevant. Mammal feces were identified by their color, shape, smell and associated footprints (Bang and Dahlstrom, 2001). Fecal and regurgitated samples were air dried at room temperature and stored individually in paper bags. Each fecal sample was later washed using a sieve under running water, and seeds were immediately and carefully removed and dried (Fedriani and Delibes, 2009).
Seed Germination Experiment
We sowed C. humilis mammal-ingested seeds as well as control seeds (i.e., non-ingested). To obtain control seeds, ripe fruits were collected during the sampling surveys from individuals distributed along the transects. Fruit ingestion and digestion have two effects on seeds that usually enhance their germination: (1) neutralization of the pulp inhibitory effect due to the removal of the pulp, and (2) seed coat scarification effect (Samuels and Levey, 2005). To separate both effects, we used two types of control seeds: intact fruits and fruits with the pulp manually removed. Therefore, we established five seed treatments for the germination experiment: (1) control 1 or “control seeds with pulp” (n = 43), (2) control 2 or “manually-depulped control seeds” (n = 44), (3) seeds regurgitated by goats (n = 99), (4) seeds spit out by goats (n = 80), and (5) pine marten-ingested seeds (n = 24). Distinction between seeds spit out and seeds regurgitated was required as the former experience a mechanical and chemical treatment only in the mouth while the latter spend also a given amount of time within the stomach in contact with the gastric juices. So, we categorized spit and regurgitated seeds based on our previous field observations and our work on other similar systems (Delibes et al., 2017a; Castañeda et al., 2018). Regurgitated seeds showed little persisting pulp, a wear out aspect, and often an irregular surface. Spit seeds have generally attached most of the pulp and a much less wear out aspect. Firstly, collected seeds were examined visually with 20–40 x magnification glasses, and traits which could affect germination, such as seed predation by insects, were recorded. Seed predation was estimated from the presence and number of holes on the endocarp (Rodríguez et al., 2014). In January 2018, both control (n = 87) and mammal-ingested seeds (n = 203) were sown individually in pots within 18 pot-trays (18 × 8 × 8 cm) with commercial substrate. Sown seeds (n = 290) were incubated in a chamber under controlled environment (25 ± 5°C, 50 ± 5% relative humidity, 12-h dark/12-h light photoperiod, uninterrupted ventilation, watering periodically; Salvador and Lloret, 1995; González-Benito et al., 2006; Hasnaoui et al., 2009). We monitored seedling emergence weekly for 9 months (from January to September), recording the date that any seedling part first emerged from the substrate surface. At the end of the experiment (33 weeks after sowing), we measured the length of emerged seedlings and extracted them and separated the aboveground part (i.e., leaves), the underground part (i.e., root), and the seed. Both aboveground and underground parts were dried for 72 h at 50°C and weighed to determine dry biomass.
Statistical Analyses
Data on seedling emergence and seedling growth were analyzed through linear mixed models using the “lme4” package in Rstudio v. 1.1.383 (R Core Team, 2017). To analyze the final proportion of seedling emergence (or emergence success), we constructed generalized linear mixed models (GLMMs) with binomial error distribution (and associated logit link). To the response variable, the value “0” was assigned to those seeds that had no emerged and “1” when it did emerge. “Seed treatment” was the only predictor variable. To analyze seedling growth (using as response variables “Seedling length” and “Seedling dry weight”), we constructed linear mixed models (LMMs) with a gaussian error distribution, using as predictor variables “Seed treatment” and “Emergence time” (i.e., number of weeks between sowing and emergence). Due to the fact that seedlings developed either one or two leaves, three linear models were constructed for the response variable “Seedling length”: model 1 with the length of the first (i.e., the highest) leaf as response variable, model 2 with the additive length (i.e., the addition of the length of the two leaves), and model 3 with the absence or presence of the second leaf as a binomial distribution. Regarding the variable response “Seedling dry weight”, we constructed three linear models as well: model 1 with the aboveground dry weight, model 2 with the underground dry weight, and model 3 with the total weight. The variable “Study site” was included as random effect in all models. We fitted the GLMMs using Laplace approximation to maximum likelihood and the LMMs using restricted maximum likelihood (REML). Adjusted means and standard errors were calculated using the package “lsmeans”, as well as the contrasts among the different levels of significant main factors through a Tukey post-hoc test.
On the other hand, to assess the potential effects of the different treatments on the emergence rate (speed), we used failure-time analyses by fitting a Cox proportional hazard regression mixed model (Therneau and Grambsch, 2000) to data consisting of the number of weeks between sowing and seedling emergence. To separate the effects on emergence rate from those on emergence success, we only considered seeds that had emerged by the end of our germination experiment (e.g., Fedriani et al., 2012). “Study site” was included as frailty factor. The significance of the target factor was evaluated by backwards stepwise elimination from the full model. In comparing successive models, we calculated the double absolute difference of their respective expectation maximization (EM) likelihood algorithms and compared that value against a chi-square with k−1 degrees of freedom, k being the number of levels of the factor being tested. For the frailty factor we also assumed a chi-square distribution with one degree of freedom (Therneau and Grambsch, 2000). For this purpose, we used the package “survival” in Rstudio v. 1.1.383 (R Core Team, 2017).
Results
Quantitative Effectiveness: Seeds per Sample and per Distance
Overall, we collected 56 mammal samples, of which 17 belonged to goat (Capra hircus L.) samples (both spits and regurgitations, Figures 3A,B) and 39 were pine marten (Martes martes L.) feces (Figure 3C). A total of 203 seeds of C. humilis were recovered from the samples: 179 seeds from goat samples, specifically 99 regurgitated seeds and 80 spit out seeds, and 24 seeds from pine marten feces. Goat samples were present in three of four study sites, with a frequency of seed occurrence of 100%. Pine marten feces were found in all study sites, although the frequency of C. humilis seed occurrence was 50% in Cala Bóquer, 25% in Ermita de Betlem and 0% in the rest of the study sites (Table S1). Therefore, the mean number of samples that contained seeds per transect was 4.25 ± 1.65 for goat and 2.25 ± 1.65 for pine marten (Figure 4A). Regarding the number of seeds per sample, it was much lower for pine marten, 2.2 ± 1.8, than for goat, 8.1 ± 3.6 (Figure 4B). The difference increases when considering the total number of seeds dispersed by each species (i.e., multiplying the number of samples with seeds by the number of seeds per sample), being 44.7 ± 19.7 for goat and 6.0 ± 3.7 for pine marten (Figure 4C). Finally, the total number of seeds dispersed corrected by the length (km) of each transect shows again that goats are much more important quantitatively (165.6 ± 138.5) than pine martens (9.5 ± 8.2) (Figure 4D). Specifically, 94.6 ± 79.8 seeds per km were regurgitated and 71.0 ± 58.8 seeds per km were spit out by goats.

Figure 3. Local mammal fruit consumers of Chamaerops humilis identified in the study. (A) A goat (Capra hircus) in front of several individuals of dwarf palm. (B) C. humilis seeds expeled by goat. (C) Pine marten (Martes martes) feces with C. humilis seeds.
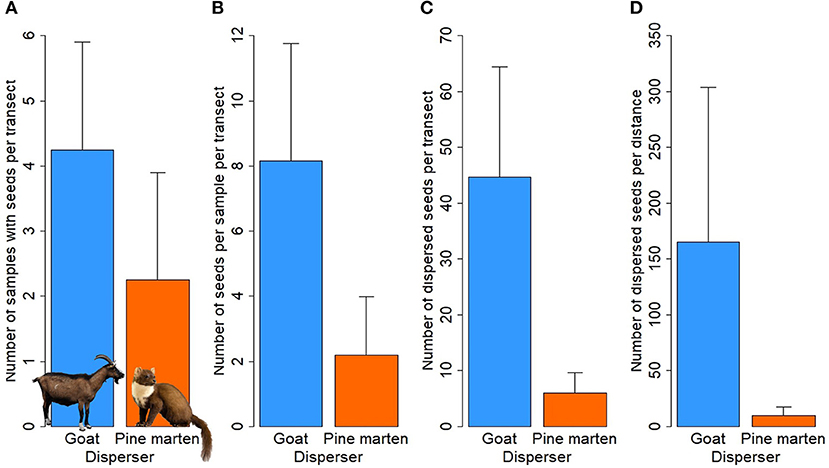
Figure 4. Means (± SE) among the four transects of different quantitative variables for the two dispersers, goat (blue bars) and pine marten (orange bars). (A) number of samples with seeds per transect; (B) number of seeds per sample per transect; (C) number of dispersed seeds per transect; (D) number of dispersed seeds per transect corrected by distance (km).
Qualitative Effectiveness: Seedling Emergence
Overall, 43.45% of the sown seeds (n = 290) germinated. Treatment-specific final emergence percentages were, on average, 56.3 ± 13.6% (n = 43), 81.1 ± 8.4% (n = 44), 27.9 ± 7.9% (n = 99), 49.4 ± 4.4% (n = 80), and 63.3 ± 3.3% (n = 24) for control seeds with pulp, manually-depulped control seeds, seeds regurgitated by goats, seeds spit out by goats, and pine marten-ingested seeds, respectively. Nonetheless, if we consider seeds regurgitated and spit out by goats together, the final emergence percentage of seeds dispersed by goats is 32.7 ± 5.7% (n = 179). Seedling emergence started 6 weeks after sowing (March 2018) for all treatments except for control seeds with pulp (C1), which started 3 weeks later (April 2018; Figure 5A). No seedling emerged in the last 8 weeks of monitoring. The Cox regression analyses indicated that, once corrected for the effect of study site (random or frailty factor), there were strong and significant differences among treatments in emergence rate (χ2 = 29.43, df = 4, P < 0.0001). Thus, on average, manually-depulped control seeds and pine marten-ingested seeds emerged 2.4 weeks earlier than control seeds with pulp. Seeds regurgitated and seeds spit out by goats emerged 5.0 and 3.3 weeks earlier than control seeds with pulp, respectively (Figure 5B). The hazard ratio of control seeds with pulp was <1 (0.4), which suggests that the presence of fruit pulp, and thus germination inhibitors, delayed seedling emergence.
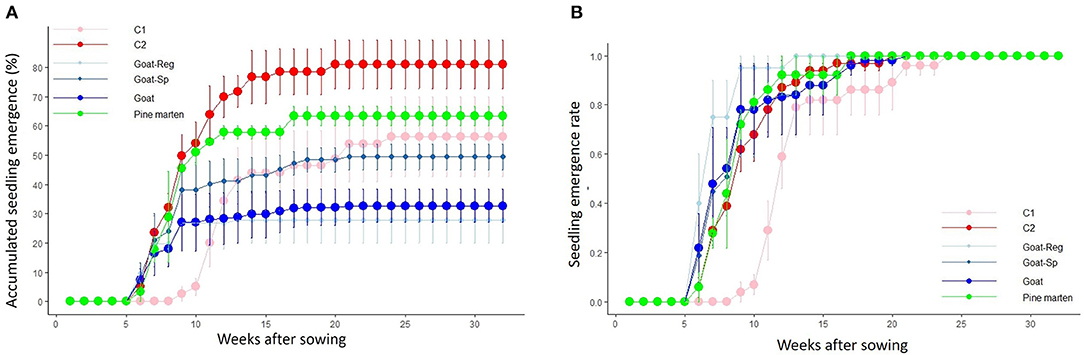
Figure 5. (A) Cumulative percentages (mean ± SE) of seedling emergence (i.e., emergence success) for the five treatments, adding the net percentages for the goat, over the weeks after sowing. (B) Emergence rate, i.e., emerged seedlings each week divided by the total emerged seedlings, for the five treatments over the weeks after sowing. Pink line, C1 or “control seeds with pulp”; red line, C2 or “manually-depulped control seeds”; light blue line, seeds regurgitated by goats; dark blue line, seeds spit out by goats; blue line, seeds dispersed by goats; and green line, pine marten-ingested seeds.
Our mixed model revealed that seed treatment had a significant effect (χ2 = 41.37, df = 4, P < 0.0001) on final emergence success (Table 2), being highest for manually-depulped control seeds and lowest for seeds regurgitated by goats (Figure 6A). In pairwise comparisons (Table S2), differences were significant between control seeds with pulp and seeds regurgitated by goats, manually-depulped control seeds and seeds regurgitated by goats, manually-depulped control seeds and seeds spit out by goats, seeds regurgitated and seeds spit out by goats, and seeds regurgitated by goats and pine marten-ingested seeds. About 24.6% of mammal dispersed seeds collected in the field (n = 203) were predated by insects (i.e., they had at least one beetle exit hole). Specifically, 18.7% (n = 80) of seeds spit out by goats and 35.3% (n = 99) of seeds regurgitated by goats had between 1 and 4 holes made by scolitine beetles, probably introduced (Rodríguez et al., 2014). No predated seeds were found for other treatments. To better understand the relative qualitative effectiveness of seed dispersers in scenarios of absence of scolitine beetles, we ran the linear mixed model again but without considering scolitine-predated seeds. Results were very similar to the previous model, again with a significant effect of seed treatment (χ2 = 28.14, df = 4, P < 0.0001) on emergence success (Table 2), but differences were significant only in pairwise comparisons between each treatment and seeds regurgitated by goats (Table S2). Adjusted means of seeds regurgitated and spit out by goats were slightly higher in this case, but without substantial changes with respect to the other treatments (Figure 6B).
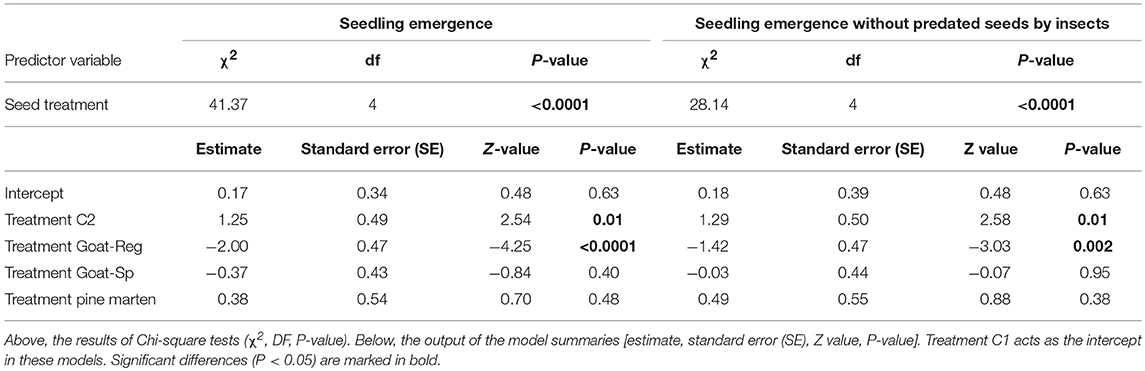
Table 2. Main results of the generalized linear mixed models testing the effects of emergence success, with and without predated seeds by insects (response variable), for each treatment (C1 or “control seeds with pulp”, C2 or “manually-depulped control seeds”, Goat-Reg or seeds regurgitated by goat, Goat-Sp or seeds spit out by goat, and Pine marten or pine marten-ingested seeds) (predictor variable).
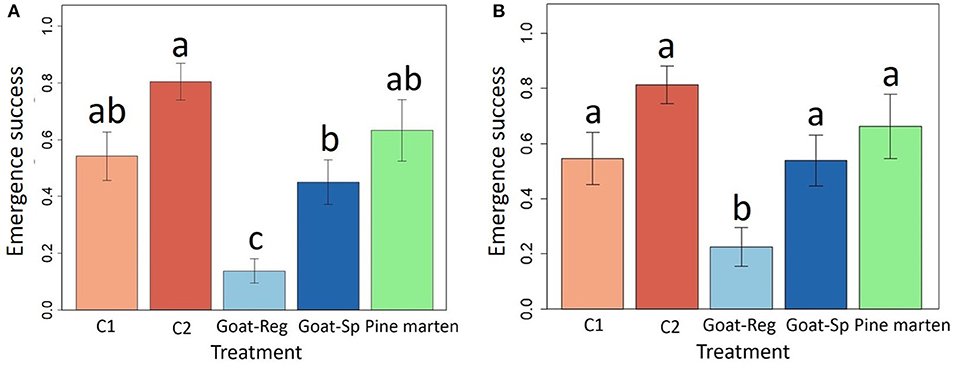
Figure 6. Model-adjusted means (± SE) of the final emergence success, with (A) and without predated seeds by insects (B), for the five treatments (C1 or “control seeds with pulp”, C2 or “manually-depulped control seeds”, Goat-Reg or seeds regurgitated by goat, Goat-Sp or seeds spit out by goat, and Pine marten or pine marten-ingested seeds). Different letters denote significant differences between paired levels in Tukey test.
Qualitative Effectiveness: Seedling Growth
Once the study site effect was corrected for, our linear mixed models revealed that seed treatment only had a marginal effect on seedling length for the first (and largest) leaf (χ2 = 8.74, df = 4, P = 0.07; Table 3). However, emergence time had a significant effect on seedling length in the three models (χ2 = 16.43, df = 1, P < 0.0001), always reaching higher values those that emerged earlier. In order to evaluate whether the relationship between emergence time and seedling length was consistent among treatments, we fitted simple linear regressions using the length of the first leaf as variable. The correlation was significantly negative for the whole dataset including all treatments and controls (P < 0.0001, Figure S1A) and for both control treatments “C1” and “C2” (P < 0.0001, Figure S1B and P < 0.05, Figure S1C, respectively). Nevertheless, it was non-significant for seeds regurgitated by goats, seeds spit out by goats, and pine marten-ingested seeds (Figures S1D–F), suggesting that seed processing by dispersers altered the negative relationship between emergence time and seedling length.
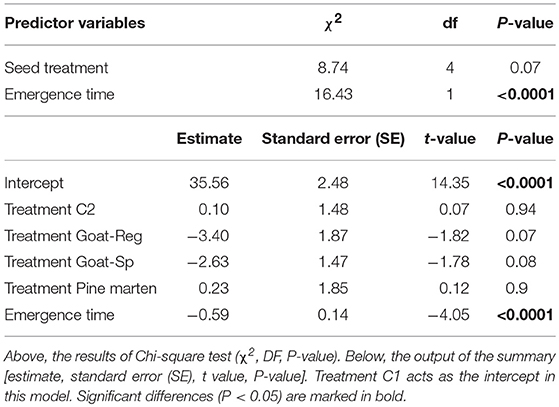
Table 3. Main results of the linear mixed model testing the effects of seed treatment (C1 or “control seeds with pulp”, C2 or “manually-depulped control seeds”, Goat-Reg or seeds regurgitated by goat, Goat-Sp or seeds spit out by goat, and Pine marten or pine marten-ingested seeds) and emergence time (predictor variables) on seedling length for the first leaf (response variable).
Seed treatment had a significant effect on seedling underground dry weight and total dry weight (χ2 = 12.94, df = 4, P = 0.01 and χ2 = 10.29, df = 4, P < 0.05, respectively), being highest for pine marten-ingested seeds and lowest for seeds spit out by goats (Figures 7A,B), but showed no effect on seedling aboveground dry weight (χ2 = 4.89, df = 4, P = 0.3; Figure 7C). In addition, emergence time had a significant effect on seedling aboveground, underground and total dry weight (P < 0.01; Table 4); seedlings emerging earlier showed higher dry weights. Pairwise comparisons through Tukey test (Table S3) indicated that differences were significant between manually-depulped control seeds—seeds spit out by goats, and seeds spit out by goats—pine marten-ingested seeds for underground dry weight. However, for aboveground dry weight differences among seed treatments were non-significant. Finally, for total dry weight, differences were only significant between manually-depulped control seeds and seeds spit out by goats.
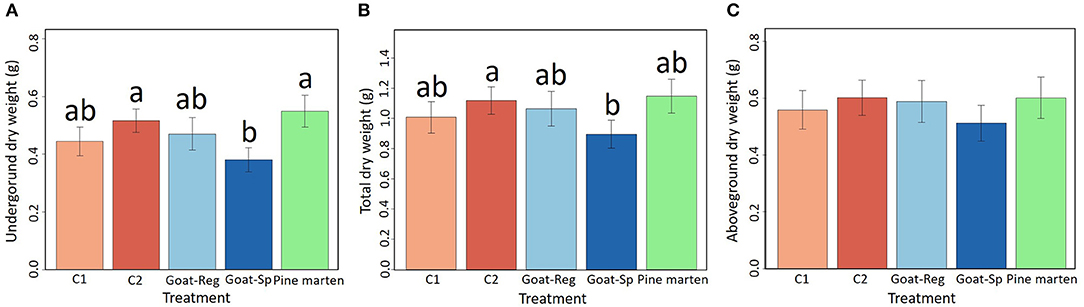
Figure 7. Model-adjusted means (± SE) of seedling underground dry weight (A), seedling total dry weight (B), and seedling aboveground dry weight (C) for the five treatments (C1 or “control seeds with pulp”, C2 or “manually-depulped control seeds”, Goat-Reg or seeds regurgitated by goat, Goat-Sp or seeds spit out by goat, and Pine marten or pine marten-ingested seeds). Different letters denote significant differences between paired levels in Tukey test.
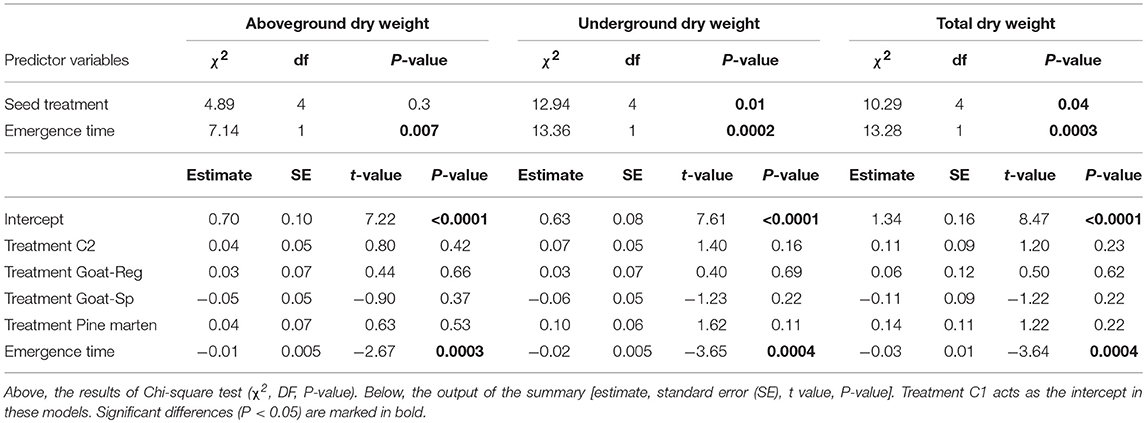
Table 4. Main results of the linear mixed models testing the effects of seed treatment (C1 or “control seeds with pulp”, C2 or “manually-depulped control seeds”, Goat-Reg or seeds regurgitated by goat, Goat-Sp or seeds spit out by goat, and Pine marten or pine marten-ingested seeds) and emergence time (predictor variables) on seedling growth: aboveground, underground, and total seedling dry weight (response variable).
Total Seed Dispersal Effectiveness (SDE)
The SDE landscape shows that goats are, overall, about nine times more effective seed dispersers (SDE = 54.14) than pine martens (SDE = 6.03) (Figure 8). Goats are high-quantity and medium-quality dispersers, whereas pine martens are high-quality but low-quantity dispersers. Moreover, if goat-treatments are separated, seeds spit out are better qualitatively (SDE = 35.07) than seeds regurgitated (SDE = 26.4). The higher overall dispersal effectiveness of goats occurred even though some goat-processed seeds were predated by scolitines.
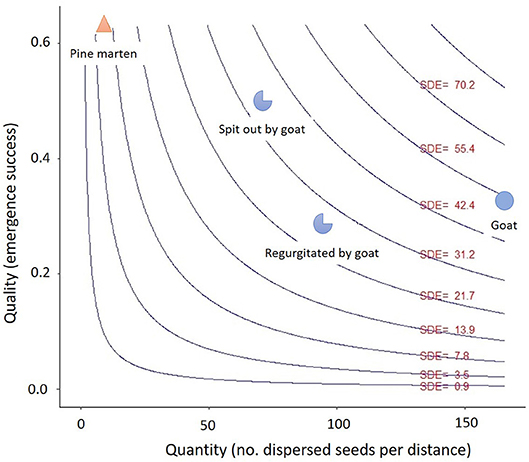
Figure 8. Seed dispersal effectiveness (SDE) landscape. The blue circles indicate the quantitative and qualitative components for goat-treatments, both separated (regurgitated and spit out seeds) and together, and the orange triangle the quantitative and qualitative components for pine marten. The isoclines (blue lines) and the SDE values (red) show the seed dispersal effectiveness (quantity x quality) throughout the SDE landscape.
Discussion
Island systems are well-known to be fauna-depauperated and, consequently, to have highly specialized plant-animal mutualisms, such as plant-frugivore interactions (e.g., González-Castro et al., 2012). Therefore, past fauna extinctions may lead to important ecological consequences for plant communities on islands (Hansen and Galetti, 2009; Traveset and Richardson, 2014), like the functional replacement of the extinct seed dispersers by contemporary frugivore assemblages (e.g., Traveset, 1995; Celedón-Neghme et al., 2013). This study assesses for the first time the SDE for C. humilis on islands, and describes understudied dispersal systems, that is, goats (C. hircus) and pine martens (M. martes) as seed dispersers of this palm in the largest Balearic island. Moreover, this study contributes with a valuable example of how non-native species can be exerting an important ecosystem function that was likely lost thousands of years ago, when humans led to the extinction of the unique native mammals on many Mediterranean islands.
Quantitative Effectiveness
Our results indicate that the quantitative component of SDE was high for goats and low for pine martens. Several variables could be explaining these large differences between both species. First, the local abundance of the disperser is a relevant variable for SDE (Schupp et al., 2010). In this case, an extremely high abundance of goats has been reported in Mallorca (it is estimated that several tens of thousands live in the island—exact amount not known; Vives and Baraza, 2010). They are heterogeneously distributed throughout Serra de Tramuntana and Serra de Llevant (Bioatles—Govern de les Illes Balears2). Second, the number of fruits handled per visit seems to be much higher for goats than for pine martens, and the handling behavior could also influence on the final number of seeds dispersed per feeding bout. Given that humans are largely determining the number of goats on Mallorca (Mayol et al., 2017), they are probably influencing the “relative seed dispersal effectiveness”, not only for C. humilis, but also for many other systems where livestock “rescues” the seed dispersal service of anachronistic plant species (Janzen and Martin, 1982; Cares et al., 2018).
Goats have been described to expel the ingested seeds while defecating (e.g., Mancilla-Leytón et al., 2011) or ruminating (e.g., Delibes et al., 2017a); however, we found no goat feces with C. humilis seeds in our study sites. On the other hand, introduced common genets (G. genetta) were scarce in the study area and no feces were found during the surveys; moreover, this species usually defecates in rather unsuitable sites for seed germination and seedling establishment (rocks and cliffs; Traveset, 2002). Likewise, we detected no signs of rabbit or rodents fruit consumption in our transects. We must note, however, that the sampling surveys were focused on forest tracks, which could be overestimating the quantitative contribution of some frugivore species—like the pine marten, which is known to use tracks to fecal marking (Barja, 2005) -, at the same time that we could have underestimated the contribution of others, like the common genet. On the other hand, pulp feeders (rabbits and rodents) could not have been identified through the fecal sampling surveys since they usually consume the pulp, leaving the seeds under the mother plant (Fedriani et al., 2012).
Qualitative Effectiveness
Seed dispersers can affect germination via two effects: (1) neutralization of the pulp inhibitory effect due to the removal of the pulp, and (2) seed coat scarification effect (Samuels and Levey, 2005). In our study, the earliest emergence of both manually-depulped control seeds and ingested seeds indicates that disperser species influence emergence time by removing the pulp inhibitory effect (Fedriani and Delibes, 2009). In relation to seed coat scarification, seed treatment in goat-stomach during rumination (i.e., digestive contractions and fermentation) could be accelerating seed germination as seed regurgitated by goats showed the highest emergence speed. However, this treatment is sometimes too harsh for seed viability, leading to the damage of seed embryos (i.e., seed predation) (e.g., Mancilla-Leytón et al., 2015; Jara-Guerrero et al., 2018) and, consequently, decreasing the final emergence success. Pine marten-ingested seeds showed the highest emergence success (after the manually-depulped control seeds); however, this disperser does not seem to be exerting any effect on emergence speed through seed scarification. Regarding seedling growth, seeds spit out by goats showed the lowest values of seedling dry weight, whereas pine marten-ingested seeds the highest ones. This could suggest that treatment in the mouth of goats could be more aggressive than treatment in the gut of pine martens, since chewing process is very intensive in ruminant species (Mancilla-Leytón et al., 2015). Nonetheless, the seedling dry weight of seeds regurgitated by goats was rather similar to that of pine marten-ingested seeds (Figures 7A,B), thus further research is required to go in depth in these results. In addition, it is necessary to consider that the method used to classify seeds regurgitated vs. seeds spit out by goats might be not entirely objective. As it is already known, seed treatment in mouth and gut differs between frugivore species –and might differ as well among conspecific individuals- as a consequence of differences in morphological and physiological traits, as well as in retention times (Traveset et al., 2007). In conclusion, the monitoring of seedling emergence and seedling growth suggests a higher qualitative seed dispersal effectiveness for pine martens than for goats.
Frugivore-plant interactions can turn out into trade-offs, i.e., benefits and costs, for plant fitness (e.g., Fedriani and Delibes, 2011). Regarding the quality of seed deposition, all the seeds predated by scolitine beetles found in the field had been dispersed by goats, specifically 18.7% (n = 80) of the seeds spit out and 35.3% (n = 99) of the seeds regurgitated. This might be related with seed covering, since pine-marten dispersed seeds are embedded in the dung (Traveset et al., 2007), but might also be due to persisting pulp surrounding the seed, which is probably higher for seeds spit out by goats and pine marten-ingested seeds than for those regurgitated by goats. Thus, pulp removal by frugivorous vertebrates appears to enhance germination, but at the same time it represents a risk due to the cost of insect predation increase (Rodríguez et al., 2014). Moreover, seeds dispersed by goats are deposited in aggregates of ~8 seeds on average, and it is very likely that high seed density has a negative effect on seedling establishment by intraspecific competition (Murray, 1998). Finally, it is well-known that the microsite of seed arrival is a key qualitative aspect of seed dispersal effectiveness (García-Cervigón et al., 2018), so it must be considered in future studies where seedling emergence and survival are monitored under field conditions.
Total Seed Dispersal Effectiveness (SDE)
Overall, the relative contribution of goats to the seed dispersal effectiveness of C. humilis is more important than that of pine martens, mainly due to the quantitative component. Goat long-distance mobility, the great amount of fruits per time that they can consume, and a long seed retention time in the gut support their functional relevance in many dispersal systems (Mancilla-Leytón et al., 2011; Grande et al., 2013). In Spain, goats have been shown to disperse effectively the seeds of at least 12 species, including C. humilis (Robles et al., 2005; Mancilla-Leytón et al., 2011, 2015; Delibes et al., 2017a). Pine martens also transport seeds over long distances (González-Varo et al., 2013; up to ~1,200 m) and have long retention times in the gut (Schaumann and Heinken, 2002). Moreover, they are known to consume many fleshy-fruited species in Northern Spain (Clevenger, 1993; Bermejo and Guitián, 1996; Rosellini et al., 2007), as well as C. humilis among other species in the Balearic Islands (Clevenger, 1995); however, the number of fruits per feeding bout is usually very low, and their population densities are not as high as those of goats.
We must consider, however, that goats not only act as effective seed dispersers, but also as herbivores, browsing frequently on C. humilis (Rivera Sánchez, 2014) and devastating leaves and inflorescences of many other plants (Mayol et al., 2017); and also as pre-dispersal seed predators (Mancilla-Leytón et al., 2011) like other ruminant species (e.g., Giordani, 2008). These ungulates have been reported to cause much damage on a number of plant species (Rivera-Sánchez et al., 2015), many of them threatened (Mayol et al., 2017). On the other hand, endemic plant species in Mallorca seem to have evolved under the selective pressure of herbivore ungulates (Vives and Baraza, 2010; Bover et al., 2016), and moderate browsing by goats indeed helps to maintain plant communities (Johnson, 2009; Rosa García et al., 2012). All these potential goat negative effects on other life plant stages of C. humilis as well as goat impact on other plant populations prevent considering them as necessarily positive from a conservation point of view.
Goat and Pine Marten: Surrogate Dispersers of Myotragus balearicus?
The extinct Balearic goat-like Myotragus balearicus Bate, the only medium-sized mammal present on the island in the Pleistocene, was a generalist species with a broad dietary niche, encompassing leaves, stems and inflorescences of diverse plant species (Bartolomé et al., 2011; Winkler et al., 2013a; Rivera et al., 2014; Welker et al., 2014). Several authors have suggested the pivotal role of this species as shaper of vegetation dynamics, acting as a keystone species in these insular ecosystems during the Pleistocene and Holocene (Palmer et al., 1999; Winkler et al., 2013b; Welker et al., 2014). Although no seeds have been found so far in any coprolites (Rivera et al., 2014; Welker et al., 2014), it is not discarded that M. balearicus could also feed on fruits (as many goats do today, e.g., Mancilla-Leytón et al., 2015; Delibes et al., 2017a) and discard seeds intact, thus acting also as a legitimate disperser. On the other hand, even though the origin of C. humilis in the Balearic Islands remains uncertain (Carrión, 2012), its old presence in the eastern coast of the Iberian Peninsula (Arroyo et al., 2004; Pérez-Obiol et al., 2010) would make more plausible the hypothesis of a Balearic existence before the recent human arrival. García-Castaño et al. (2014) even suggest that Balearic dwarf palm populations might appeared because of microplate migrations from the Oligocene or during the Messinian salinity crisis. In addition, an active human transport of C. humilis should have taken place along the Mediterranean basis in the Quaternary (García-Castaño et al., 2014; Guzmán et al., 2017).
Our study demonstrates that both goats and pine martens are effective seed dispersers of C. humilis in Mallorca, and we thus hypothesize that these introduced frugivores, especially the goat, could be replacing the function probably carried out by the extinct M. balearicus in the past, acquiring the role of seed dispersal “rescuers”. This role has already been suggested for the goat in other systems (e.g., Mancilla-Leytón et al., 2015; Cares et al., 2018). So, despite the fitness costs imposed by both introduced mammals, they could be providing an indispensable service to this ancient palm by dispersing it on island systems, where native dispersers are extinct and the alternatives are very limited.
Conclusions
In spite of the myriad negative impacts widely described for non-native species on islands (Reaser et al., 2007; Mayol et al., 2017; Rogers et al., 2017; Weller et al., 2018), their role as rescuers of ecological functions, lost in the past with fauna extinctions, acquires great relevance under the current scenario of increasing defaunation (Dirzo et al., 2014; Malhi et al., 2016). Our results here support the Janzen and Martin (1982) anachronism hypothesis and suggest that non-native species can disperse effectively anachronistic fruits, exerting a fundamental role in insular ecosystems. Thus, more studies are required to assess the role of non-native species in depauperated communities and to establish suitable management programs to preserve plant species and their associated ecological functions in Mediterranean ecosystems.
Author Contributions
JMF and AT: experimental design and field work. RM-G and JMF: laboratory work. RM-G: statistical analyses and original paper draft. RM-G, JMF, and AT: paper review and editing.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to J. A. Alcover, E. Baraza, and J. Mayol for information about the current situation of C. hircus on Mallorca island, to F. Burjachs and J. Rita for solving some doubts about the C. humilis pollen dating in the Balearics, and to J. P. González-Varo for contributing to the last paper review. Thanks also to S. Santamaría for helping with the field work. This study is framed within project CGL2017-88122-P financed by the Spanish Government. JMF was funded by Portuguese Foundation for Science and Technology (grant IF/00728/2013).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00161/full#supplementary-material
Footnotes
References
Alcover, J. A. (1984). Über die nahrung der ginsterkatze genetta genetta (Linnaeus, 1758) auf den inseln mallorca, ibiza und cabrera. Säugetierkundliche Mitt. 31, 189–195.
Alcover, J. A., Sans, A., and Palmer, M. (1998). The extent of extinctions of mammals on islands. J. Biogeogr. 25, 913–918. doi: 10.1046/j.1365-2699.1998.00246.x
Arroyo, J., Carrión, J., Hampe, A., and Jordano, P. (2004). “La distribución de las especies a diferentes escalas espacio-temporales,” in Ecología del Bosque Mediterráneo En Un Mundo Cambiante, ed F. Valladares (Madrid: Ministerio de Medio Ambiente, EGRAF), 27–67.
Barja, I. (2005). Patrones de marcaje con heces por la marta europea (Martes martes) en el noroeste de España: importancia para su estudio. Galemys 17, 123–134.
Bartolomé, J., Retuerto, C., Martínez, X., Alcover, J. A., Bover, P., Cassinello, J., et al. (2011). Sobre la dieta de Myotragus balearicus, un bóvido sumamente modificado del Pleistoceno-Holoceno de las Baleares. Nat. Aragon. 27, 4–7.
Beaune, D., Fruth, B., Bollache, L., Hohmann, G., and Bretagnolle, F. (2013). Doom of the elephant-dependent trees in a Congo tropical forest. For. Ecol. Manage. 295, 109–117. doi: 10.1016/j.foreco.2012.12.041
Bello, C., Galetti, M., Pizo, M. A., Magnago, L. F. S., Rocha, M. F., Lima, R. A. F., et al. (2015). Defaunation affects carbon storage in tropical forests. Sci. Adv. 1:e1501105. doi: 10.1126/sciadv.1501105
Bermejo, T., and Guitián, J. (1996). Consumo de frutos y dispersión de semillas de serbal (Sorbus aucuparia L.) por zorros y martas en la Cordillera cantábrica occidental. Doñana Acta Vertebr. 23, 215–227.
Biondi, E., and Filigheddu, R. (1990). A palm fossil closely related to Chamaerops humilis L. from the lower miocene of sardinia. G. Bot. Ital. 124, 711–724. doi: 10.1080/11263509009428134
Bond, W. J., Lee, W. G., and Craine, J. M. (2004). Plant structural defences against browsing birds: a legacy of New Zealand's extinct moas. Oikos 104, 500–508. doi: 10.1111/j.0030-1299.2004.12720.x
Bover, P., and Alcover, J. A. (2008). Extinction of the autochthonous small mammals of Mallorca (Gymnesic Islands, Western Mediterranean) and its ecological consequences. J. Biogeogr. 35, 1112–1122. doi: 10.1111/j.1365-2699.2007.01839.x
Bover, P., Quintana, J., and Alcover, J. A. (2008). Three islands, three worlds: paleogeography and evolution of the vertebrate fauna from the Balearic Islands. Quat. Int. 182, 135–144. doi: 10.1016/j.quaint.2007.06.039
Bover, P., Valenzuela, A., Torres, E., Cooper, A., Pons, J., and Alcover, J. A. (2016). Closing the gap: new data on the last documented Myotragus and the first human evidence on Mallorca (Balearic Islands, Western Mediterranean Sea). Holocene 26, 1887–1891. doi: 10.1177/0959683616645945
Bueno, R. S., Guevara, R., Ribeiro, M. C., Culot, L., Bufalo, F. S., and Galetti, M. (2013). Functional redundancy and complementarities of seed dispersal by the last neotropical megafrugivores. PLoS ONE 8:e56252. doi: 10.1371/journal.pone.0056252
Cancio, I., González-Robles, A., Bastida, J. M., Isla, J., Manzaneda, A. J., Salido, T., et al. (2017). Landscape degradation affects red fox (Vulpes vulpes) diet and its ecosystem services in the threatened Ziziphus lotus scrubland habitats of semiarid Spain. J. Arid Environ. 145, 24–34. doi: 10.1016/j.jaridenv.2017.05.004
Cares, R. A., Sáez-Cordovez, C., Valiente-Banuet, A., Medel, R., and Botto-Mahan, C. (2018). Frugivory and seed dispersal in the endemic cactus Eulychnia acida: extending the anachronism hypothesis to the Chilean Mediterranean ecosystem. Rev. Chil. Hist. Nat. 91:9. doi: 10.1186/s40693-018-0079-4
Carrión, J. S. (2002). A taphonomic study of modern pollen assemblages from dung and surface sediments in arid environments of Spain. Rev. Palaeobot. Palynol. 120, 217–232. doi: 10.1016/S0034-6667(02)00073-8
Carrión, J. S. (2012). Paleoflora y Paleovegetación de la Península Ibérica e Islas Baleares: Plioceno-Cuaternario, 1st Edn. Murcia: Ministerio de Economía y Competitividad.
Castañeda, I., Fedriani, J. M., and Delibes, M. (2018). Potential of red deer (Cervus elaphus) to disperse viable seeds by spitting them from the cud. Mamm. Biol. 90, 89–91. doi: 10.1016/j.mambio.2017.10.004
Caughlin, T. T., Ferguson, J. M., Lichstein, J. W., Zuidema, P. A., Bunyavejchewin, S., and Levey, D. J. (2015). Loss of animal seed dispersal increases extinction risk in a tropical tree species due to pervasive negative density dependence across life stages. Proc. R. Soc. B Biol. Sci. 282, 1–9. doi: 10.1098/rspb.2014.2095
Ceballos, G., Ehrlich, P. R., Barnosky, A. D., García, A., Pringle, R. M., and Palmer, T. M. (2015). Accelerated modern human-induced species losses: entering the sixth mass extinction. Sci. Adv. 1:e1400253. doi: 10.1126/sciadv.1400253
Celedón-Neghme, C., Traveset, A., and Calviño-Cancela, M. (2013). Contrasting patterns of seed dispersal between alien mammals and native lizards in a declining plant species. Plant Ecol. 214, 657–667. doi: 10.1007/s11258-013-0197-7
Clevenger, A. P. (1993). Pine marten (Martes martes Linné, 1758) comparative feeding ecology in an island and mainland population of Spain. Z. Säugetierkd. 58, 212–224.
Clevenger, A. P. (1995). Seasonality and relationships of food resource use of martes martes, genetta genetta and felis catus in the Balearic Islands. Rev. d'Ecol. Terre Vie 50, 109–131.
Clout, M., and Hay, J. (1989). The importance of birds as browsers, pollinators and seed dispersers in New Zealand forests. N. Z. J. Ecol. 12, 27–33.
Collevatti, R. G., Grattapaglia, D., and Hay, J. D. (2003). Evidences for multiple maternal lineages of Caryocar brasiliense populations in the Brazilian Cerrado based on the analysis of chloroplast DNA sequences and microsatellite haplotype variation. Mol. Ecol. 12, 105–115. doi: 10.1046/j.1365-294X.2003.01701.x
Delibes, M., Castañeda, I., and Fedriani, J. M. (2017a). Tree-climbing goats disperse seeds during rumination. Front. Ecol. Environ. 15, 222–223. doi: 10.1002/fee.1488
Delibes, M., Centeno-Cuadros, A., Muxart, V., Delibes, G., Ramos-Fernández, J., and Morales, A. (2017b). New insights into the introduction of the common genet, Genetta genetta (L.) in Europe. Archaeol. Anthropol. Sci. 2017, 1–9. doi: 10.1007/s12520-017-0548-8
Dirzo, R., Young, H. S., Galetti, M., Ceballos, G., Isaac, N. J. B., and Collen, B. (2014). Defaunation in the antrhopocene. Science 345, 401–406. doi: 10.1126/science.1251817
Eddine, A., Mostefai, N., Smet, K., De Klees, D., Ansorge, H., Karssene, Y., et al. (2017). Diet composition of a newly recognized canid species, the African Golden Wolf (Canis anthus), in Northern Algeria. Ann. Zool. Fenn. 54, 347–356. doi: 10.5735/086.054.0506
El Alami, A., and Chait, A. (2017). Etude de l'alimentation du magot Macaca sylvanus dans le site touristique des cascades d'Ouzoud (Maroc). Revue de Primatologie [En ligne], 7 | 2016, mis en ligne le 22 février 2017. doi: 10.4000/primatologie.2748
Farris, E., Canopoli, L., Cucca, E., Landi, S., Maccioni, A., and Filigheddu, R. (2017). Foxes provide a direct dispersal service to Phoenician junipers in mediterranean coastal environments: ecological and evolutionary implications. Plant Ecol. Evol. 150, 117–128. doi: 10.5091/plecevo.2017.1277
Faurby, S., and Svenning, J. C. (2015). Historic and prehistoric human-driven extinctions have reshaped global mammal diversity patterns. Divers. Distrib. 21, 1155–1166. doi: 10.1111/ddi.12369
Fedriani, J. M. (1996). Dieta anual del zorro, Vulpes vulpes, en dos hábitats del Parque Nacional de Doñana. Doñana Acta Vertebr. 23, 143–152.
Fedriani, J. M., and Delibes, M. (2008). ¿Quien siembra los arbustos en Doñana? Un estudio sobre dispersión de semillas por mamíferos. Quercus 271, 23–28.
Fedriani, J. M., and Delibes, M. (2009). Functional diversity in fruit-frugivore interactions : a field experiment with Mediterranean mammals. Ecography 32, 983–992. doi: 10.1111/j.1600-0587.2009.05925.x
Fedriani, J. M., and Delibes, M. (2011). Dangerous liaisons disperse the Mediterranean dwarf palm: fleshy-pulp defensive role against seed predators. Ecology 92, 304–315. doi: 10.1890/09-2194.1
Fedriani, J. M., Zywiec, M., and Delibes, M. (2012). Thieves or mutualists? Pulp feeders enhance endozoochore local recruitment. Ecology 93, 575–587. doi: 10.1890/11-0429.1
Galetti, M., Donatti, C. I., Pires, A. S., Guimarães, P. R., and Jordano, P. (2006). Seed survival and dispersal of an endemic Atlantic forest palm: the combined effects of defaunation and forest fragmentation. Bot. J. Linn. Soc. 151, 141–149. doi: 10.1111/j.1095-8339.2006.00529.x
Galetti, M., Guevara, R., Côrtes, M. C., Fadini, R., Matter, S., Von Leite, A. B., et al. (2013). Functional extinction of birds drives rapid evolutionary changes in seed size. Sci. Rep. 340, 1086–1090. doi: 10.1126/science.1233774
Galetti, M., Moleón, M., Jordano, P., Pires, M. M., Guimarães, P. R., Pape, T., et al. (2018). Ecological and evolutionary legacy of megafauna extinctions. Biol. Rev. 93, 845–862. doi: 10.1111/brv.12374
Garcia, C., and Morey, M. (1989). Distribución geográfica de la vegetación de matorral en la zona occidental de Mallorca. Treballs Geogr. 41, 33–46.
García-Castaño, J. L., Terrab, A., Ortiz, M. Á., Stuessy, T. F., and Talavera, S. (2014). Patterns of phylogeography and vicariance of Chamaerops humilis L. (Palmae). Turk. J. Botany 38, 1132–1146. doi: 10.3906/bot-1404-38
García-Cervigón, A. I., Zywiec, M., Delibes, M., Suárez-Esteban, A., Perea, R., and Fedriani, J. M. (2018). Microsites of seed arrival: spatio-temporal variations in complex seed-disperser networks. Oikos 127, 1001–1013. doi: 10.1111/oik.04881
Giordani, L. (2008). The role of goats in germination and dispersal of mimosa luisana brandegee (leguminosae-mimosoideae) seeds in tehuacan-cuicatlan Valley, Puebla State, Mexico. (master thesis). Department of International Environment and Development Studies; Norwegian University of Life Sciences (UMB).
González-Benito, M. E., Huertas-Micó, M., and Pérez-García, F. (2006). Seed germination and storage of Chamaerops humilis (dwarf fan palm). Seed Sci. Technol. 34, 143–150. doi: 10.15258/sst.2006.34.1.15
González-Castro, A., Traveset, A., and Nogales, M. (2012). Seed dispersal interactions in the Mediterranean Region: contrasting patterns between islands and mainland. J. Biogeogr. 39, 1938–1947. doi: 10.1111/j.1365-2699.2012.02693.x
González-Varo, J. P., López-Bao, J. V., and Guitián, J. (2013). Functional diversity among seed dispersal kernels generated by carnivorous mammals. J. Anim. Ecol. 82, 562–571. doi: 10.1111/1365-2656.12024
Grande, D., Mancilla-Leytón, J. M., Delgado-Pertiñez, M., and Martin-Vicente, A. (2013). Endozoochorus seed dispersal by goats: recovery, germinability and emergence of five Mediterranean shrub species. Spanish J. Agric. Res. 11, 347–355. doi: 10.5424/sjar/2013112-3673
Guimarães, P. R., Galetti, M., and Jordano, P. (2008). Seed dispersal anachronisms: rethinking the fruits extinct megafauna ate. PLoS ONE 3:e9307. doi: 10.1371/journal.pone.0001745
Guzmán, B., Fedriani, J. M., Delibes, M., and Vargas, P. (2017). The colonization history of the Mediterranean dwarf palm (Chamaerops humilis L., Palmae). Tree Genet. Genom. 13, 1–10. doi: 10.1007/s11295-017-1108-1
Hansen, D. M., and Galetti, M. (2009). The forgotten megafauna. Science 324, 42–43. doi: 10.1126/science.1172393
Hasnaoui, O., Bouazza, M., and Thinon, M. (2009). Kinetical germination study of the Chamaerops humilis L. var. argentea Andre (Arecaceae). Environ. Res. J. 3, 76–80.
Herrera, J. (1989). On the reproductive biology of the dwarf palm, Chamaerops humilis in southern Spain. Principes 33, 27–32.
Jansen, P. A., Hirsch, B. T., Emsens, W.-J., Zamora-Gutierrez, V., Wikelski, M., and Kays, R. (2012). Thieving rodents as substitute dispersers of megafaunal seeds. Proc. Natl. Acad. Sci. U.S.A. 109, 12610–12615. doi: 10.1073/pnas.1205184109
Janzen, D. H. (1982). Differential seed survival and passage rates in cows and horses, surrogate Pleistocene dispersal agents. Oikos 38, 150–156. doi: 10.2307/3544014
Janzen, D. H., and Martin, P. S. (1982). Neotropical anachronisms: the fruits the gomphotheres ate. Science 215, 19–27. doi: 10.1126/science.215.4528.19
Jara-Guerrero, A., Escribano-Avila, G., Espinosa, C. I., De la Cruz, M., and Méndez, M. (2018). White-tailed deer as the last megafauna dispersing seeds in Neotropical dry forests: the role of fruit and seed traits. Biotropica 50, 169–177. doi: 10.1111/btp.12507
Johnson, C. N. (2009). Ecological consequences of late quaternary extinctions of megafauna. Proc. R. Soc. B Biol. Sci. 276, 2509–2519. doi: 10.1098/rspb.2008.1921
Jordano, P. (2014). An R Package for Plots of Effectiveness Landscapes in Mutualisms: effect.lndscp. Available online at: https://github.com/pedroj/effectiveness_pckg
Levey, D. J., Silva, W. R., and Galetti, M. (2002). “Seed dispersal and frugivory: ecology, evolution and conservation,” in eds D. J. Levey, W. R. Silva, and M. Galetti (Wallingford: CABI Publishing), 544.
Malhi, Y., Doughty, C. E., Galetti, M., Smith, F. A., Svenning, J.-C., and Terborgh, J. W. (2016). Megafauna and ecosystem function from the Pleistocene to the Anthropocene. Proc. Natl. Acad. Sci. U.S.A. 113, 838–846. doi: 10.1073/pnas.1502540113
Mancilla-Leytón, J. M., Fernandez-Alés, R., and Martín-Vicente, Á. (2015). Efectividad cualitativa del ganado caprino en la dispersión de especies de frutos secos y carnosos. Ecosistemas 24, 22–27. doi: 10.7818/ECOS.2015.24-3.04
Mancilla-Leytón, J. M., Fernández-Alés, R., and Vicente, A. M. (2011). Plant-ungulate interaction: goat gut passage effect on survival and germination of Mediterranean shrub seeds. J. Veg. Sci. 22, 1031–1037. doi: 10.1111/j.1654-1103.2011.01325.x
Martín, M. (2008). “Caracterització de la dieta de la guineu (Vulpes vulpes) al Parc del Garraf,” in V Trobada d'Estudiosos del Garraf, eds J. Hernández, J. Grau, and J. Melero (Barcelona: Diputació de Barcelona), 81–88.
Mayol, J., Alcover, J. A., Domènech, O., Moragues, E., and Rita, J. (2017). La Cabra, Espècie Invasora a les Balears. Panor. les Illes Balear. 1–96.
Mayol, J., Álvarez, C., and Manzano, X. (2009). Presència i control del coatí, Nasua nasua L., i d'altres carnívors introduïts en època recent a Mallorca. Bolleti la Soc. d'Historia Nat. Les Balear. 52, 183–191.
McConkey, K. R., and Drake, D. R. (2006). Flying foxes cease to function as seed dispersers long before they become rare. Ecology 87, 271–276. doi: 10.1890/05-0386
Medel, R., Aizen, M. A., and Zamora, R. (2009). Ecología y Evolución de Interacciones Planta-Animal, 1a Edn. Santiago de Chile: Editorial Universitaria.
Meehan, H. J., McConkey, K. R., and Drake, D. R. (2002). Potential disruptions to seed dispersal mutualisms in Tonga, Western Polynesia. J. Biogeogr. 29, 695–712. doi: 10.1046/j.1365-2699.2002.00718.x
Moncunill-Solé, B., Jordana, X., and Köhler, M. (2016). How common is gigantism in insular fossil shrews? Examining the ‘Island Rule' in soricids (Mammalia: Soricomorpha) from Mediterranean Islands using new body mass estimation models. Zool. J. Linn. Soc. 178, 163–182. doi: 10.1111/zoj.12399
Moncunill-Solé, B., Jordana, X., Marín-Moratalla, N., Moyà-Solà, S., and Köhler, M. (2014). How large are the extinct giant insular rodents? New body mass estimations from teeth and bones. Integr. Zool. 9, 197–212. doi: 10.1111/1749-4877.12063
Murray, B. R. (1998). Density-dependent germination and the role of seed leachate. Aust. J. Ecol. 23, 411–418. doi: 10.1111/j.1442-9993.1998.tb00748.x
Palmer, M., Pons, G. X., Cambefort, I., and Alcover, J. A. (1999). Historical processes and environmental factors as determinants of inter-island differences in endemic faunas: the case of the Balearic Islands. J. Biogeogr. 26, 813–823. doi: 10.1046/j.1365-2699.1999.00319.x
Palomares, F., and Delibes, M. (1991). Dieta del Meloncillo, Herpestes ichneumon, en el Coto del Rey (Norte del Parque Nacional de Doñana, S.O. de España). Acta Vertebr. 18, 187–194.
Palombo, M. R., Kohler, M., Moya Sola, S., and Giovinazzo, C. (2008). Brain versus body mass in endemic ruminant artiodactyls: a case studied of Myotragus balearicus and smallest Candiacervus species from Mediterranean Islands. Quat. Int. 182, 160–183. doi: 10.1016/j.quaint.2007.08.037
Perea, R., Delibes, M., Polko, M., Suárez-Esteban, A., and Fedriani, J. M. (2013). Context-dependent fruit-frugivore interactions: partner identities and spatio-temporal variations. Oikos 122, 943–951. doi: 10.1111/j.1600-0706.2012.20940.x
Pérez-Méndez, N., Jordano, P., García, C., and Valido, A. (2016). The signatures of Anthropocene defaunation: cascading effects of the seed dispersal collapse. Sci. Rep. 6:24820. doi: 10.1038/srep24820
Pérez-Obiol, R., Jalut, G., Julià, R., Pèlachs, A., Iriarte, M. J., Otto, T., et al. (2010). Mid-Holocene vegetation and climatic history of the Iberian Peninsula. Holocene 21, 75–93. doi: 10.1177/0959683610384161
Pinna, W., Nieddu, G., Moniello, G., and Cappai, M. G. (2007). Vegetable and animal food sorts found in the gastric content of Sardinian Wild Boar (Sus scrofa meridionalis). J. Anim. Physiol. Anim. Nutr. 91, 252–255. doi: 10.1111/j.1439-0396.2007.00700.x
Pinya, S., Perelló, E., and Àlvarez, C. (2009). Sobre la presencia del mapache Procyon lotor (Linnaeus, 1758) en la isla de Mallorca. Galemys 21, 61–64.
Pires, M. M., Guimarães, P. R., Galetti, M., and Jordano, P. (2018). Pleistocene megafaunal extinctions and the functional loss of long-distance seed- dispersal services. Ecography 41, 153–163. doi: 10.1111/ecog.03163
R Core Team (2017). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online at: https://www.R-project.org/
Reaser, J. K., Meyerson, L. A., Cronk, Q., De Poorter, M., Eldrege, L. G., Green, E., et al. (2007). Ecological and socioeconomic impacts of invasive alien species in island ecosystems. Environ. Conserv. 34, 98–111. doi: 10.1017/S0376892907003815
Requena-Mullor, J. M., López, E., Castro, A. J., Virgós, E., and Castro, H. (2016). Landscape influence on the feeding habits of European badger (Meles meles) in arid Spain. Mammal Res. 61, 197–207. doi: 10.1007/s13364-016-0269-x
Revilla, E., and Palomares, F. (2002). Does local feeding specialization exist in Eurasian badgers? Can. J. Zool. 80, 83–93. doi: 10.1139/z01-208
Ripple, W. J., Newsome, T. M., Wolf, C., Dirzo, R., Everatt, K. T., Galetti, M., et al. (2015). Collapse of the world's largest herbivores. Sci. Adv. 1:e1400103. doi: 10.1126/sciadv.1400103
Rivera Sánchez, L. (2014). Ecología Trófica De Ungulados En Condiciones De Insularidad. Available online at: https://hdl.handle.net/10803/285367
Rivera, L., Baraza, E., Alcover, J. A., Bover, P., Rovira, C. M., and Bartolomé, J. (2014). Stomatal density and stomatal index of fossil Buxus from coprolites of extinct Myotragus balearicus Bate (Artiodactyla, Caprinae) as evidence of increased CO2 concentration during the late Holocene. Holocene 24, 876–880. doi: 10.1177/0959683614530445
Rivera-Sánchez, L., Cassinello Roldán, J., Baraza Ruíz, E., and Bartolomé Filellla, J. (2015). Comparative study of trophic behaviour and herd structure in wild and feral goats living in a mediterranean island: management implications. Appl. Anim. Behav. Sci. 165, 81–87. doi: 10.1016/j.applanim.2015.01.015
Robles, A. B., Castro, J., and Ramos, M. E. (2005). “Effects of ruminal incubation and goats' ingestion on seed germination of two legume shrubs: adenocarpus decorticans Boiss. and Retama sphaerocarpa (L.) Boiss.,” in Options Méditerranéennes: Série A. Séminaires Méditerranéens, eds E. M. Alcaide, H. Ben Salem, K. Biala, and P. Morand-Fehr (Zaragoza: CIHEAM), 111–115.
Rodríguez, M., Delibes, M., and Fedriani, J. M. (2014). Hierarchical levels of seed predation variation by introduced beetles on an endemic mediterranean palm. PLoS ONE 9:e109867. doi: 10.1371/journal.pone.0109867
Rogers, H. S., Buhle, E. R., HilleRisLambers, J., Fricke, E. C., Miller, R. H., and Tewksbury, J. J. (2017). Effects of an invasive predator cascade to plants via mutualism disruption. Nat. Commun. 8:14557. doi: 10.1038/ncomms14557
Rosa García, R., Celaya, R., García, U., and Osoro, K. (2012). Goat grazing, its interactions with other herbivores and biodiversity conservation issues. Small Rumin. Res. 107, 49–64. doi: 10.1016/j.smallrumres.2012.03.021
Rosellini, S., Barja, I., and Piñeiro, A. (2007). Distribución y hábitos alimenticios de la marta (Martes martes) en el parque natural os montes do invernadeiro (Galicia, NO de España). Galemys 19, 99–114.
Rotllàn-Puig, X., and Traveset, A. (2016). Declining relict plants: climate effect or seed dispersal disruption? A landscape-scale approach. Basic Appl. Ecol. 17, 81–91. doi: 10.1016/j.baae.2015.08.003
Rumeu, B., Devoto, M., Traveset, A., Olesen, J. M., Vargas, P., Nogales, M., et al. (2017). Predicting the consequences of disperser extinction: richness matters the most when abundance is low. Funct. Ecol. 31, 1910–1920. doi: 10.1111/1365-2435.12897
Salvador, R., and Lloret, F. (1995). Germinación en el laboratorio de varias especies arbustivas mediterráneas: efecto de la temperatura. Orsis 10, 25–34.
Samuels, I. A., and Levey, D. J. (2005). Effects of gut passage on seed germination: do experiments answer the questions they ask? Funct. Ecol. 19, 365–368. doi: 10.1111/j.1365-2435.2005.00973.x
Schaumann, F., and Heinken, T. (2002). Endozoochorous seed dispersal by martens (Martes foina, M. martes) in two woodland habitats. Flora 197, 370–378. doi: 10.1078/0367-2530-00053
Schupp, E. W. (1993). Quantity, quality and the effectiveness of seed dispersal by animals. Vegetatio 107/108, 15–29. doi: 10.1007/978-94-011-1749-4_2
Schupp, E. W., Jordano, P., and Gómez, J. M. (2010). Seed dispersal effectiveness revisited: a conceptual review. N. Phytol. 188, 333–353. doi: 10.1111/j.1469-8137.2010.03402.x
Schurr, M. R., Fuentes, A., Luecke, E., Cortes, J., and Shaw, E. (2012). Intergroup variation in stable isotope ratios reflects anthropogenic impact on the Barbary macaques (Macaca sylvanus) of Gibraltar. Primates 53, 31–40. doi: 10.1007/s10329-011-0268-0
Smith, F. A., Doughty, C. E., Malhi, Y., Svenning, J. C., and Terborgh, J. (2016). Megafauna in the Earth system. Ecography 39, 99–108. doi: 10.1111/ecog.02156
Sodhi, N. S., Brook, B. W., and Bradshaw, C. J. A. (2012). “Causes and consequences of species extinctions,” in The Princeton Guide to Ecology, ed S. A. Levin (Princeton, NJ: Princeton University Press), 514–520. doi: 10.1515/9781400833023.514
Suárez-Esteban, A., Delibes, M., and Fedriani, J. M. (2013). Barriers or corridors? The overlooked role of unpaved roads in endozoochorous seed dispersal. J. Appl. Ecol. 50, 767–774. doi: 10.1111/1365-2664.12080
Therneau, T. M., and Grambsch, P. M. (2000). Modeling Survival data: Extending the Cox Model, 1st Edn., eds K. Dietz, M. Gail, K. Krickeberg, J. Samet, and A. Tsiatis (New York, NY: Springer Science+Business Media, LLC), 39–78. doi: 10.1007/978-1-4757-3294-8
Thompson, J. D. (2005). Plant Evolution in the Mediterranean. New York, NY: Oxford University Press.
Traveset, A. (1995). Seed dispersal of Cneorum tricoccon L. (Cneoraceae) by lizards and mammals in the Balearic islands. Acta Oecologica 16, 171–178.
Traveset, A. (2002). Consecuencias de la ruptura de mutualismos planta-animal para la distribución de especies vegetales en las Islas Baleares. Rev. Chil. Hist. Nat. 75, 117–126. doi: 10.4067/S0716-078X2002000100011
Traveset, A., González-Varo, J. P., and Valido, A. (2012). Long-term demographic consequences of a seed dispersal disruption. Proc. R. Soc. B Biol. Sci. 279, 3298–3303. doi: 10.1098/rspb.2012.0535
Traveset, A., and Richardson, D. M. (2014). Mutualistic interactions and biological invasions. Annu. Rev. Ecol. Evol. Syst. 45, 89–113. doi: 10.1146/annurev-ecolsys-120213-091857
Traveset, A., Robertson, A. W., and Rodríguez-Pérez, J. (2007). “A review on the role of endozoochory in seed germination,” in Seed Dispersal: Theory and its Application in a Changing World, eds A. J. Dennis, E. W. Schupp, R. J. Green, and D. A. Westcott (Wallingford: CAB International), 78–103. doi: 10.1079/9781845931650.0078
Valenzuela, A., and Alcover, J. A. (2013a). Documenting introductions: the earliest evidence for the presence of dog (Canis familiaris Linnaeus 1758) in the Prehistory of the Balearic Islands. J. Isl. Coast. Archaeol. 8, 422–435. doi: 10.1080/15564894.2013.810677
Valenzuela, A., and Alcover, J. A. (2013b). Radiocarbon evidence for a prehistoric deliberate translocation: the weasel (Mustela nivalis) of Mallorca. Biol. Invasions 15, 717–722. doi: 10.1007/s10530-012-0321-y
Valenzuela, A., and Alcover, J. A. (2015). The chronology of the introduction of two species of Martes (Carnivora, Mustelidae) on the Western Mediterranean Islands: first direct radiocarbon evidence. Biol. Invasions 17, 3093–3100. doi: 10.1007/s10530-015-0947-7
Valiente-Banuet, A., Aizen, M. A., Alcántara, J. M., Arroyo, J., Cocucci, A., Galetti, M., et al. (2015). Beyond species loss: the extinction of ecological interactions in a changing world. Funct. Ecol. 29, 299–307. doi: 10.1111/1365-2435.12356
Vives, J. A., and Baraza, E. (2010). La cabra doméstica asilvestrada (Capra hircus) en Mallorca ¿Una especie a erradicar? Galemys 22, 193–205.
Welker, F., Duijm, E., van der Gaag, K. J., van Geel, B., de Knijff, P., van Leeuwen, J., et al. (2014). Analysis of coprolites from the extinct mountain goat Myotragus balearicus. Quat. Res. 106–116. doi: 10.1016/j.yqres.2013.10.006
Weller, S. G., Sakai, A. K., Clark, M., Lorence, D. H., Flynn, T., Kishida, W., et al. (2018). The effects of introduced ungulates on native and alien plant species in an island ecosystem: implications for change in a diverse mesic forest in the Hawaiian Islands. For. Ecol. Manage. 409, 518–526. doi: 10.1016/j.foreco.2017.11.023
Winkler, D. E., Schulz, E., Calandra, I., Gailer, J. P., Landwehr, C., and Kaiser, T. M. (2013a). Indications for a dietary change in the extinct Bovid genus Myotragus (Plio-Holocene, Mallorca, Spain). Geobios 46, 143–150. doi: 10.1016/j.geobios.2012.10.010
Winkler, D. E., van den Hoek Ostende, L. W., Schulz, E., Calandra, I., Gailer, J. P., Landwehr, C., et al. (2013b). Dietary divergence in space and time - Lessons from the dwarf-goat Myotragus balearicus (Pleisto-Holocene, Mallorca, Spain). Mamm. Biol. 78, 430–437. doi: 10.1016/j.mambio.2013.08.003
Wood, J. R., Rawlence, N. J., Rogers, G. M., Austin, J. J., Worthy, T. H., and Cooper, A. (2008). Coprolite deposits reveal the diet and ecology of the extinct New Zealand megaherbivore moa (Aves, Dinornithiformes). Quat. Sci. Rev. 27, 2593–2602. doi: 10.1016/j.quascirev.2008.09.019
Wotton, D. M., and Kelly, D. (2011). Frugivore loss limits recruitment of large-seeded trees. Proc. R. Soc. B 278, 3345–3354. doi: 10.1098/rspb.2011.0185
Keywords: anachronism, goat, megafauna extinction, Myotragus balearicus, pine marten, seed dispersal effectiveness, seedling emergence, seedling growth
Citation: Muñoz-Gallego R, Fedriani JM and Traveset A (2019) Non-native Mammals Are the Main Seed Dispersers of the Ancient Mediterranean Palm Chamaerops humilis L. in the Balearic Islands: Rescuers of a Lost Seed Dispersal Service? Front. Ecol. Evol. 7:161. doi: 10.3389/fevo.2019.00161
Received: 31 October 2018; Accepted: 23 April 2019;
Published: 24 May 2019.
Edited by:
Casper H. A. Van Leeuwen, Netherlands Institute of Ecology (NIOO-KNAW), NetherlandsReviewed by:
Carl R. Gosper, Department of Biodiversity, Conservation and Attractions (DBCA), AustraliaAlejandro Valenzuela, University of Barcelona, Spain
Copyright © 2019 Muñoz-Gallego, Fedriani and Traveset. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raquel Muñoz-Gallego, raquel19026@gmail.com