 Erin K. Buchholtz1,2,3,4*
Erin K. Buchholtz1,2,3,4* Lauren Redmore2,4,5
Lauren Redmore2,4,5 Lee A. Fitzgerald1,2,3
Lee A. Fitzgerald1,2,3 Amanda Stronza2,4,5
Amanda Stronza2,4,5 Anna Songhurst4,6Graham McCulloch4,6
Anna Songhurst4,6Graham McCulloch4,6- 1Ecology and Evolutionary Biology Program, Texas A&M University, College Station, TX, United States
- 2Applied Biodiversity Science Program, Texas A&M University, College Station, TX, United States
- 3Department of Wildlife and Fisheries Sciences, Biodiversity Research and Teaching Collections, Texas A&M University, College Station, TX, United States
- 4Ecoexist Project, Maun, Botswana
- 5Department of Recreation, Parks, and Tourism Sciences, Texas A&M University, College Station, TX, United States
- 6Department of Zoology, University of Oxford, Oxford, United Kingdom
In social-ecological systems around the world, human-wildlife interactions are on the rise, often with negative consequences. This problem is particularly salient in areas where populations of humans and wildlife are increasing and share limited space and resources. However, few studies look at how both people and wildlife navigate shared spaces. To better examine people and wildlife within the same environment, we used methods from social science and spatial ecology to investigate how humans and elephants in Botswana utilize trees, a shared natural resource. Trees provide an opportunity to study shared resource use because they are important for people as firewood and for elephants as food and habitat. We compared tree species gathered on 49 firewood collections with the species damaged by elephants in 83 vegetation plots. We found that many tree species were damaged by elephants in ways that would generate firewood. There was also a strong overlap in the tree species that people collected and the species that elephants browsed and/or damaged. We compared spatially-explicit firewood collection locations and movement data from elephant GPS collars to model resource selection by people and elephants. Proximity to settlements was a strong driving factor for people in firewood collection, while various factors including vegetation characteristics played a role in predicting elephant movement. We found that areas where people collect firewood were negatively correlated with daytime elephant movement and positively correlated with nighttime elephant movement. We further compared the times that people collected firewood with the times when elephants were near the villages and found that people collected firewood during daylight hours when elephants were not nearby, providing further evidence of temporal partitioning. People and elephants utilized the same species of trees, and also had correlated spatial patterns of resource selection. Therefore, elephant foraging of trees provides a previously unrecognized utility to people in the form of firewood creation, and temporal partitioning allows this to occur without direct human-elephant interaction.
Introduction
In many parts of the world, humans and wildlife increasingly share land and resources outside of protected areas. Studies on human-wildlife interactions within social-ecological systems (SES) often focus on competition for resources or direct loss caused to one species by the other. These interactions are typically examined through the lens of human-wildlife conflict. Studies that focus solely on conflict between wildlife and humans may overlook other possible kinds of interactions, including benefits that one species may provide to the other. As proposed frameworks for coexistence focus on balancing inputs of costs and benefits (Kansky et al., 2016; Ceauşu et al., 2019), overlooking certain interactions can lead to incomplete understanding of the system and an overestimation of conflict. Research that focuses on the diversity of interactions, including benefits, can improve the understanding of the complexity of human-wildlife relations, including conditions for coexistence (Frank, 2016).
There is no species that exemplifies the challenges of human-wildlife conflict like the elephant. Where populations of people and elephants overlap, interactions increase and so does the potential for conflict situations (Sitati et al., 2003; Graham et al., 2010). The problem of crop loss and property damage caused by elephants is well studied (e.g., Spinage, 1990; Naughton-Treves, 1997; Sitati et al., 2005; Pozo et al., 2017). Elephants are dangerous and injury and death from elephant encounters is not rare. In southern Africa, reports show that elephants were responsible for the deaths of 31 people in Mozambique from July 2006 through September 2008 (Dunham et al., 2010), and five people per year in South Africa (Twine and Magome, 2007). In our study site in the Eastern Panhandle of the Okavango Delta, Botswana, elephants are responsible for at least one human death per year (personal observation). Encounters that don't result in human death or injury may result in significant psychological stress that carries other social costs (Ogra, 2008; DeMotts and Hoon, 2012; Mayberry et al., 2017), including decreased tolerance for elephants (Mariki et al., 2015). Human-elephant conflict (HEC) threatens the lives and livelihoods of the people who live with elephants, and is an existential threat to the long-term survival of elephant populations where they remain today.
In places where people live alongside elephants, efforts to reduce impacts of interactions have relied on a variety of mitigation measures that bear significant costs. As examples, farmers may relocate to guard their fields against elephant foraging during the growing season, may change their daily activities to avoid encounters, forgo gathering resources in areas frequented by elephants, or devote money and time to building fences and implementing deterrents (Mayberry et al., 2017). People may even kill elephants in self-defense or in retaliation (Obanda et al., 2008; Roever et al., 2013). HEC can also have wide-ranging impacts on elephants, specifically their movements and resource-use. For example, elephants often change their behavior by moving in larger groups (Songhurst et al., 2016), moving faster, and altering daily activity patterns (Graham et al., 2009). In studies looking at human-elephant interactions within SES, studies have focused separately on the behaviors of people or elephants (e.g., Sitati et al., 2003; Ogra, 2008; Graham et al., 2010; Jadhav and Barua, 2012; Mackenzie and Ahabyona, 2012; Osipova et al., 2018). This leaves many questions unanswered about the nature of human-elephant relations and provides a compelling gap in knowledge for interdisciplinary research and synthesis.
Trees provide an opportunity to study the demands for shared natural resources by elephants and people. Elephants depend on trees for protection, shade for thermoregulation, and are an important source of food. People depend on the wood from trees for firewood and construction materials for canoes and homes, as well as fruits that can help buffer food insecurity (Mmopelwa et al., 2009). Most HEC research to date has focused on direct and indirect conflict around private resources, such as property damage and agricultural crop-raiding. The use of shared common pool resources, in this case trees, is an important component of human-elephant interactions that remains to be explored. In particular, elephants browse and break trees, creating a source of accessible, dead wood that is readily collected by people for firewood.
We integrated approaches from landscape ecology and anthropology to investigate how people and elephants use tree resources in a shared landscape. We hypothesized that because elephants damage trees, which creates downed wood we would find a pattern of resource overlap among the tree species damaged by elephants and the tree species that people rely on for firewood. We also hypothesized that we would find positively correlated spatial patterns of resource use by people and elephants, but temporal resource use would differ.
Firewood and Rural Development
For most of human history, people have relied on woodfuel, including firewood and charcoal, for energy (Goren-Inbar et al., 2000). Woodfuel remains one of the most important sources of energy for rural subsistence households who often access it for free (Leach and Mearns, 1988; Dewees, 1989). In rural places with limited economic opportunities, subsistence households may depend solely on firewood for cooking, heating water, for light in the evening, and other uses that help to buffer the effects of poverty and underdevelopment (Dewees, 1989; Shackleton et al., 2007).
Beginning in the mid-1970s, scholars sounded the alarm over concerns of the “fuelwood gap” (Eckholm, 1975). Rising prices of fossil fuels led scientists to fear that a global setback in energy diversification, in particular for growing populations in developing countries, would increase the environmental pressure on forests for fuelwood production (Arnold et al., 2003). It was assumed that this rebounding demand for woodfuel may lead to a fuelwood shortage, ultimately increasing rates of deforestation (Eckholm, 1975). However, anticipated rates of deforestation fell short of projections, in large part because firewood use depends on a suite of variables, including global economies, rural-urban migration trends, rural development, and cultural preferences, among other factors (Dovie et al., 2004; Cline-Cole, 2007; Hiemstra-van der Horst and Hovorka, 2009). Studies now show that rural firewood supply is often met through a combination of waste wood generated during land clearing and harvesting of building poles, in addition to collecting fallen, dead branches, or by hacking off live branches from trees, a practice akin to coppicing that in some cases may increase woodland productivity (Dewees, 1989). The amount of firewood that subsistence households can harvest is further constrained by both the availability of individuals to contribute labor, as well as the firewood weight that people can physically carry on their heads or shoulders (Agarwal, 1986).
Studies that observed local or regional-level forest degradation linked to fuelwood harvest have concluded that degradation is due to the intersection of diverse factors, including socio-economic factors like rapid population growth, changing labor supplies, and global markets, as well as environmental factors like low tree density and climactic conditions (Arnold et al., 2006; Mills Busa, 2013; Shaheen et al., 2016). When firewood becomes scarce due to demand that outpaces environmental capacity, rural households will employ a suite of coping strategies, including traveling longer distances to harvest dead wood, harvesting live trees, cooking less frequently or cooking food of lower nutritional value, and cooking with extended family to pool firewood resources (Leach and Mearns, 1988; Dewees, 1989). Ecological studies conducted in temperate and tropical forests show that the removal of dead wood influences biodiversity through fire regimes, nutrient cycling, seed establishment, and food and habitat resources for insects, fungi, and bacteria (Jonsson et al., 2005; Cornwell et al., 2009; Joseph et al., 2011; Bouget et al., 2012; Brais and Drouin, 2012; Jacobs and Work, 2012; Olou et al., 2019). The ecological impacts of harvesting dead wood from semi-arid environments remain largely unknown (Shackleton, 1998) but may have no significant impact on vegetation (Vázquez et al., 2011).
Elephant Diet and Impact on Woody Vegetation
Elephants are generalist megaherbivores, and shrubs and trees make up a significant portion of their diet depending on availability (Cerling et al., 2006; Woolley et al., 2009; Owen-Smith and Chafota, 2012; Shrader et al., 2012). Elephants can subsist on relatively low quality food (Sukumar, 2003; Owen-Smith and Chafota, 2012) and have rapid digestive throughput which allows them to take advantage of fibrous woody plants to meet their nutritional demands (Boundja and Midgley, 2010). In addition to being valuable as food resources, trees represent a critical component of the habitat for savanna elephants. One major eco-physiological consideration for elephants is temperature. Due to their thermal inertia it is difficult for elephants to dissipate heat and they may prefer wooded habitat, which offers shade (Kinahan et al., 2007; Mole et al., 2016). Elephants may also select wooded habitats for protection or refuge when they are stressed or exposed to anthropogenic disturbance (Jachowski et al., 2013).
As a consequence of elephants seeking out wooded habitat for foraging, shade, and refuge, they in turn influence that habitat. Extensive research has been done on the effects of elephants on trees; their influence on physical structure, growth, and community composition has been documented throughout Africa (Laws, 1970; Jachmann and Croes, 1991; Ben-Shahar, 1993; Calenge et al., 2002; Skarpe et al., 2004; Guldemond and Van Aarde, 2008; Ihwagi et al., 2010). Notably, elephant damage can create a coppicing effect (Lewis, 1991; Sheil and Salim, 2004; O'Connor et al., 2007). Although severe structural damage to the main trunk or uprooting may result in tree mortality, coppicing allows trees to regenerate branches from around the damaged base and survive (Stokke and du Toit, 2000; O'Connor et al., 2007; Vanak et al., 2012). These direct effects on trees will then affect the ecosystem in varied ways, and the impact is often so apparent and widespread that elephants are considered ecosystem engineers (Jones et al., 1994; Haynes, 2012). For example, the increased structural complexity of elephant-damaged trees and elephant-modified habitats has been linked to higher levels of herpetofaunal richness (Nasseri et al., 2011) and higher densities of lizards (Pringle, 2008). Elephant activity and impact can also influence a variety of processes from nutrient cycling to community composition of herbivore guilds (Fritz et al., 2002; Skarpe et al., 2004).
A significant by-product of elephant damage to trees is in the generation of downed wood. As mentioned above, this coarse woody debris serves an ecosystem function of their own (Jonsson et al., 2005). In arid rangelands, dead shrubs may not decompose for over a decade (Milton and Dean, 1996), leaving behind woody debris in the environment. However, further research is needed to understand the role of downed branches and other woody debris within the Okavango ecosystem. Our study did not aim to quantify amounts of dead wood generated by elephants nor the amount collected by people, although these will likely have consequences on ecosystem processes. Our work here is focused on the ways that people may benefit from elephant modification of woodlands and how humans and elephants may reduce their direct interactions. Our findings bear important implications for understanding and mitigating human-elephant conflict.
Methods
Study Site
The study site is in the Eastern Panhandle of the Okavango Delta (Panhandle), Botswana, by one of the largest intact wetlands in the world (Figure 1). The Okavango Delta is formed from the Okavango River and floods seasonally, with an inland alluvial fan approximately 40,000 km2 in size (Gumbricht et al., 2004). The Okavango Delta in Botswana is a Ramsar Wetland of International Importance and UNESCO World Heritage Site, supplied by year-round water from the Okavango River that sustains both people and wildlife. The Panhandle is bordered by the national boundary fence with Namibia to the north, and the national Northern Buffalo veterinary cordon fence to the east. The region is a semi-arid savanna, with dominant vegetation classes of thornbush savannas, woodlands on Kalahari sand, shrublands, and the wetlands associated with the Delta. Approximately 18,000 African elephants (Loxodonta africana) currently range through the Eastern Panhandle where they compete for scarce resources with roughly 16,000 people (Ecoexist, 2017; Pozo et al., 2017). It is crucial to understand that the elephant population in this region is sizeable, while human development is sparse.
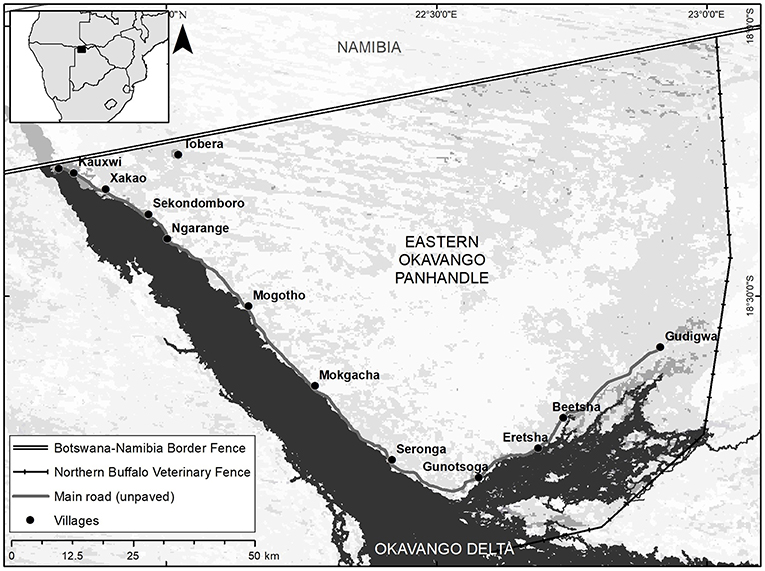
Figure 1. Map of the study region in the Eastern Okavango Panhandle, Botswana.
People in the Panhandle live across 14 designated villages and many unofficial settlements. Villages range from a population of 475 people in Tobera to a population of 3,716 people in Seronga, the sub-district capital and the largest village in the study site (Botswana Central Statistics Office, 2011). In the Panhandle, firewood is the most important form of domestic energy (Mmopelwa et al., 2009). Rural households have limited economic opportunities and electricity or bottled cooking gas are largely unaffordable or unavailable. Mmopelwa et al. (2009) report that households in the Panhandle prefer 20 species of tree for firewood, all of which have little to no financial value within the study site. While properties such as burn rate are important in determining the efficiency of different species of wood (Tietema et al., 1991), people often choose species for firewood based on availability or personal preference rather than burning properties (Tietema et al., 1991; Tietema, 1993; Kgathi and Mlotshwa, 1997). People in the Panhandle also make decisions about which species not to collect based on cultural taboos, especially those that link the burning of particular species to livestock infertility or death, as well as other practical reasons, such as the kind of smoke produced that may irritate eyes or lungs of people in proximity to the fire.
Firewood collection is labor intensive and depends on factors such as availability, household size, and season, as people use more firewood during cold winters (Gaye, 2007). Women generally carry limited tools to assist in firewood harvest, sometimes carrying cushioning and straps, and other times stripping bark from young saplings to tie the bundle. Other people may choose to carry axes to cut larger branches or stems into manageable sizes. People often carry firewood on their heads or shoulders, but carts and motorized transport are also used, though mainly by men (Kgathi and Mlotshwa, 1997). People collect firewood near agricultural homes (Kgathi and Mlotshwa, 1997) and cattleposts when work is combined, for example when men harvest firewood on their way home from tending to livestock. Researchers, government officials, and Botswana policy documents continue to link firewood harvest to deforestation (National Council on Population and Development, 1997; Mapaure and Ndeinoma, 2011). However, in situ evidence and previous studies show that people collect dried, dead wood from the ground rather than felling live trees (Mmopelwa et al., 2009).
Mokgacha, a village in the Panhandle, was the main site for our ethnographic and vegetation fieldwork. Mokgacha is situated between two major elephant pathways, time-worn paths that elephants use to move between the savanna and the Delta's waters (Songhurst et al., 2016). Mokgacha has a population of 496 people representing BaHambukushu, BaYeyi, Boga Khwe, and Xani Khwe ethnic groups (Botswana Central Statistics Office, 2011). Mokgacha was designated as a village in 2013 and is situated along the permanent water source of the Okavango River. Mokgacha was selected because, unlike the other villages along the Okavango River, it is currently not electrified and residents are exclusively dependent on firewood for cooking food, smoking out mosquitoes during the rainy season, and evening light for entertainment or study.
People
We selected households from across Mokgacha where individuals were recruited for repeated firewood harvest focal follows (hereafter referred to as focal follows) (n = 14) (Alvard, 1993). We selected households based on certain criteria, including representation of ethnicity, residence location, and willingness to participate in the study. These factors were important as they influence how someone harvests firewood based on mode of transport, cultural taboos around certain tree species that should not be burned, and availability of certain species in proximity to residence location and harvest site. We used the household as the main unit of study because although certain individuals are most often responsible for firewood harvest within households, illness or absence meant that sometimes a different household member was responsible for firewood harvest. We conducted 49 focal follows in total from February 4, 2017 through June 15, 2018, with an average of 4 focal follows per household (see Supplementary Information).
Following verbal consent with each participant, we arranged focal follows up to a week prior to actual harvest. We met participants at their home at their preferred time and date. On some occasions participants were unavailable at the agreed upon time and focal follows were postponed to a later time and/or date. During focal follows, we used a handheld GPS unit to record both the primary collection site as well as the track traveled from the residence to reach that point. We recorded starting locations and times as well as the time that firewood harvest began and ended. We identified and listed all firewood species harvested during each focal follow. Species of firewood harvested were identified with the help of a research assistant from the locality with extensive experience collecting and identifying firewood. We recorded names in Setswana or other local languages when necessary and translated local tree names into Latin names with the assistance of Vogel (unpublished data) and Okavango Research Institute Herbarium.
Elephants
We collected data on trees and assessed elephant-related damage in vegetation plots around Mokgacha. Because we wanted a representative sample which included the variety of habitat types in the area, we chose not to assign points within a grid but instead to stratify sampling by habitat type and distance to the village. We first identified eight broad habitat types based on an existing vegetation classification (GeoTerra Image Ltd, 2007). The classes were as follows: (1) open grassland; (2) open/sparse bushland; (3) open woodland/bushland; (4) closed canopy woodland; (5) tall closed canopy forest; (6) wetland floodplain; (7) wetland seepage/pan; and (8) non-wet bare. We sampled each habitat type within four distance buffers from the edge of the village (0–250, 250–500, 500–750, and 750–1,000 m) based on a priori knowledge of firewood collection behavior. This resulted in 32 possible combinations of habitat and distance from village, of which 25 existed. We used ArcGIS 10.3 to assign 5 random points within each of these combinations with a minimum of 15 meters distance between each point. If points fell in misclassified areas (i.e., in water, fields, or roads), we relocated 10 meters into the nearest wooded habitat. If the available habitat was too small to allow 5 points, each 15 meters apart, we reduced the number of points so as not to over-sample. This led to a total of 102 vegetation plots sampled.
For each of the assigned points, we assessed a 5-meter radius plot. We recorded each tree or shrub >1 meter in height within the plot and assessed elephant damage to each individual plant. Types of elephant damage recorded included: presence/absence of browse (leaves and twigs < 2 cm diameter), presence/absence of small branch damage (2–10 cm diameter), presence/absence of large branch damage (> 10 cm diameter), presence/absence of main stem damage of any size, mortality, and percentage of uprooting and debarking around the circumference. Elephant damage was distinguished from human or livestock damage by assessing height of browsing, type of branch break or browse, manually broken tree trunks, visual hatchet marks, or low, clipped vegetation associated with cattle and goats.
We quantified how firewood would be generated by elephant damage to trees based on an acceptance-availability calculation for each species. We based our firewood generation index on a plant selection index described by Owen-Smith and Chafota (2012) for quantifying elephant foraging. We calculated the value for tree species acceptance as the number of plots where that species incurred firewood-generating damage by elephants divided by the number of plots where that species was present. We calculated each tree species' availability as the number of plots that species was present in, divided by the total number of plots. The firewood generation index for each tree species represents the species availability divided by acceptance.
We used location data from 10 male and 10 female elephants in the Panhandle to estimate spatial and temporal use of tree resources by elephants. The elephants were fitted with Vectronic GPS collars which recorded hourly location fixes (https://www.vectronic-aerospace.com/wildlife-monitoring/gps-collars). They were deployed by the Ecoexist Project (www.ecoexistproject.org) in April 2014 and removed in April 2018 with permission from the Government of Botswana under research permit reference EWT 8/36/4 XVII (79) and Immobilization permit 2014 WP/RES 15/2/2 XXIII (169). Elephants were immobilized using Thianil (thiafentanil oxalate), 15 mg per male elephant and 11–12 mg per female elephant. Darts were fired from a modified.22-caliber rifle and the effect of the immobilizing drug was reversed using intravenous Trexonil (naltrexone hydrochloride) at a dose of 10 mg for each 1 mg of Thianil. Once the elephant was recumbent, it was fitted with a satellite collar around the neck (see Songhurst, 2014 for details). We filtered all tracking data for spurious GPS fixes and error readings.
Analyses
Tree Species Use
We tabulated the tree species collected on each of the focal follows with Mokgacha households and calculated the frequency that each species was collected. We compared this frequency with the firewood generation index value calculated for each tree species that was found within the vegetation plots. We excluded plots on Delta islands for the comparison, as people were unable to reach those islands to collect firewood during our study period.
Temporal Proximity
We extracted the times that people harvested firewood from the focal follows. We calculated temporal patterns of elephant proximity to settlements by selecting all points that were within 250 m of the edge of a settlement (7,009 points). We calculated the proportion of those points that occurred during each hour of the day and compared them to the proportion of times that people collected firewood.
Spatial Resource Use
We chose to use a resource-selection approach to estimate spatially explicit probabilities of use for areas where people collect firewood. The point-based resource selection function compares resources for sampled used and available points (Boyce et al., 2002; Manly et al., 2002). The locations where people collected firewood during focal follows represented the used sample points. To represent the area available for firewood collection, we buffered Mokgacha by the average distance traveled to collect firewood joined with a minimum convex polygon based on all the points where firewood had been collected during focal follows. We then generated two random points for every true firewood collection location point within that polygon in order to represent the available area while not creating a highly zero-inflated regression. We used remotely sensed data for the variables to include in the general linear model to test likelihood of use, including Normalized Difference Vegetation Index (NDVI), vegetation class, and distance to the settlement, main road, and Delta. We chose NDVI as a proxy for vegetation presence and a classification to represent different types of vegetation. We selected distance to the settlement, main road, and Delta because these seemed to be relevant to traveling to collect resources. Slope is negligible in this region. These variables and the remote sensing methods by which they were derived are outlined in Table 1. Upon spatial assessment we removed three habitat variables as each class represented < 2% of the study area. We fitted a generalized linear model to the data using the lme4 package (Bates et al., 2015) and this and all further analyses were carried out using R (R Core Team, 2018). We used Akaike's Information Criterion (AIC) to assess our full model compared with models where each variable was removed (Akaike, 1987). We excluded the distance to Delta variable in our final model to reduce AIC. All other variables chosen a priori were included in the model because their removal represented only minor reductions in AIC and were still ecologically meaningful for our analysis.
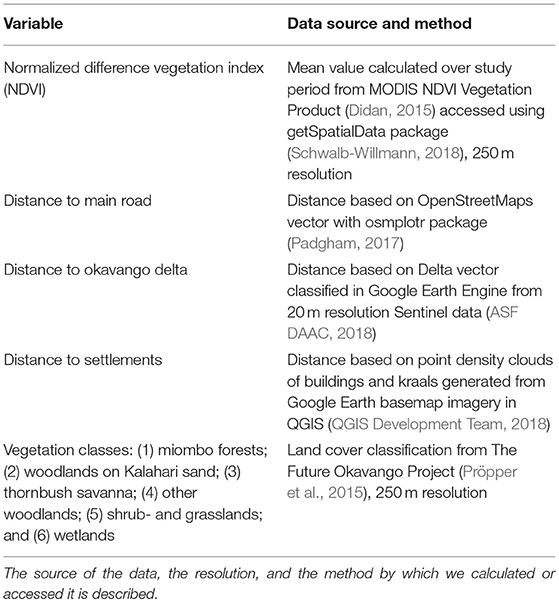
Table 1. We choose the covariates below to include in our spatial models.
We used the same point-based resource selection function method to model spatial resource use by elephants. We subsampled the points to a minimum of 4 h interval to reduce autocorrelation since the resource selection function assumes independence between points, and serial animal locations are not independent of each other. To represent the points available to elephants we generated two random points for every used point within the minimum convex polygon for each individual elephant using the amt package (Signer et al., 2018). We calculated a generalized linear model, again using the lme4 package in R (Bates et al., 2015). We used the same data sources for the covariates as for the firewood collection model (Table 1), this time including all habitats as they were widely present within the area of elephant movement. Building on our temporal analysis, we modeled elephant resource selection during the day and at night. We used AIC for model selection (Akaike, 1987), however AIC was not significantly reduced by removing any variables, and we included all a priori variables in the model.
We generated spatial prediction rasters based on the models and compared the outputs using Spearman's correlation coefficient and with the spatialEco package (Evans and Ram, 2018). We would expect to find a positive correlation coefficient if there is a similar spatial pattern of likelihood where people collect firewood and where elephants move, and the correlation raster will visualize the spatial differences in correlation.
Results
Tree Species Use
Thirty-five tree species were collected by people for firewood at least once during focal follows, and 15 were harvested on at least five or more focal follows. The vegetation plots we assessed had an average of 6 different tree species and 18 individual trees. On average, 13 trees per plot showed signs of elephant browsing, and 10 were damaged by elephants in some way that would generate firewood. Nineteen tree species occurred in the vegetation plots and elephant spoor was found in all 83 plots surveyed. Elephants damaged all 19 of the tree species to some degree, and 11 were damaged in at least 80% of the plots where they were present (firewood generation index >0.80). Of the top 10 species damaged most frequently by elephants, 8 of those species were also in the top 10 frequently collected for firewood (Figure 2). The most abundant species, Dichrostachys cinerea, was found in 62 plots and had firewood-generating damage in 52 of those plots. Sixty percent of the 230 individual D. cinerea trees we assessed had elephant damage. The most abundant tree species with a high overlap in both elephant and human use was Combretum collinum. We assessed 100 individual C. collinum trees in 30 plots, of which 90% were damaged by elephants.
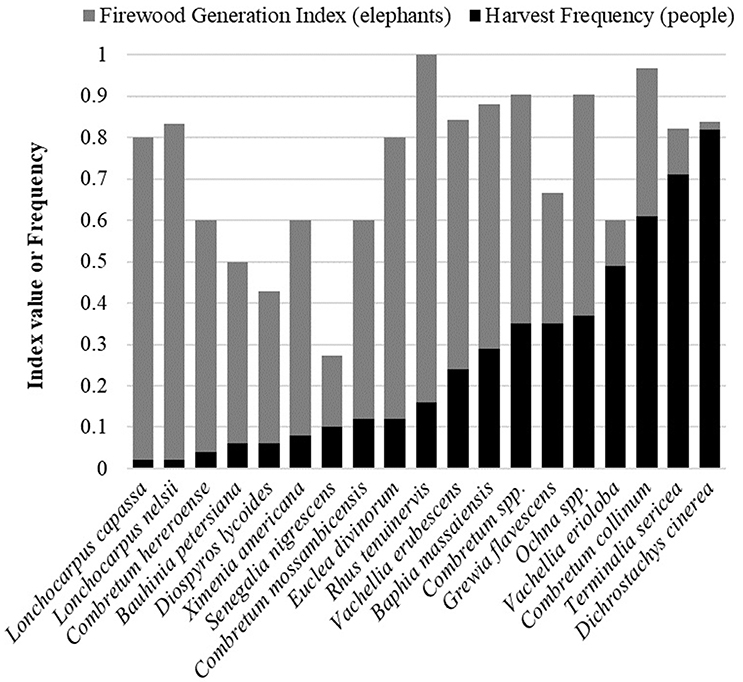
Figure 2. Plot of the frequency of firewood collection by people and the elephant firewood generation index value for each tree species. All tree species found in at least five vegetation plots were included. Species are ordered by increasing frequency of firewood harvest by people.
Temporal Proximity
We found contrasting temporal patterns of landscape use by people and elephants (Figure 3). People collected firewood most often during mid-morning and mid-afternoon. Elephants were near to settlements (< 250 m) most often at night.
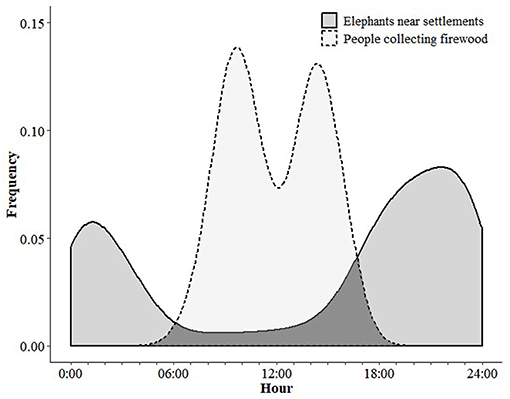
Figure 3. Contrasting temporal pattern of firewood collection by people and frequency of elephant proximity near settlements (within 250 meters).
Spatial Resource Use
Proximity to settlements was the only significant predictor of human resource selection for firewood (z = −3.324, p < 0.001). People selected for areas that were close to settlements. NDVI and habitat classes were not significant (Table 2 and Figure 4).
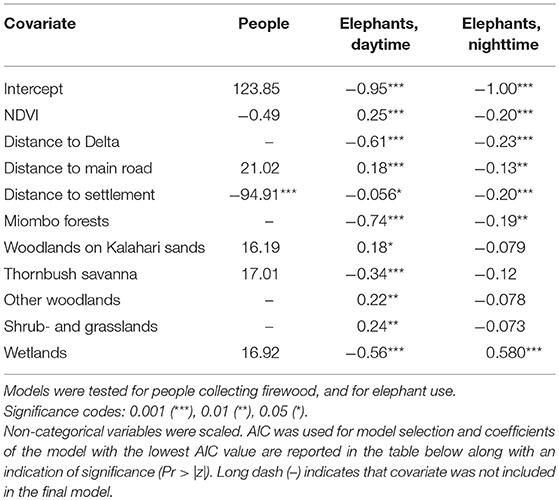
Table 2. Resource selection functions were modeled using generalized linear models.
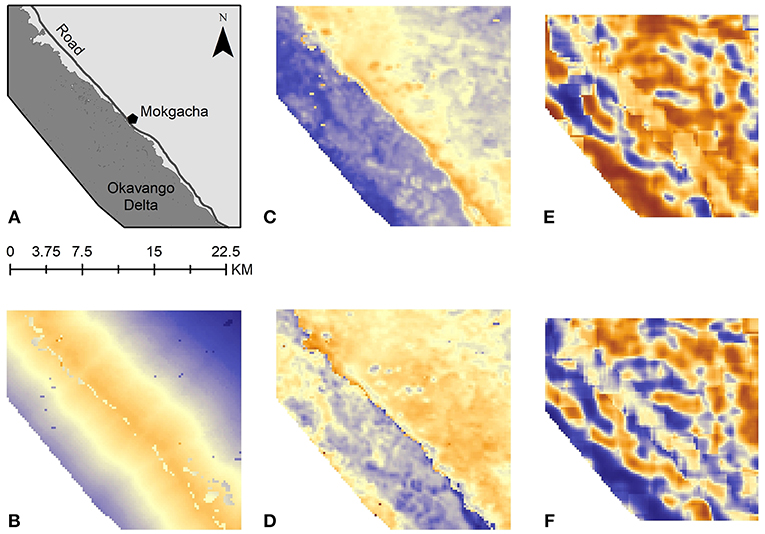
Figure 4. Predicted surfaces generated from resource selection models. The color ramp scales from negative selection values in blue to positive selection values in orange. Coefficients and covariates used to create these surfaces are reported in Table 2. (A) Reference key for region and scale, showing location of Mokgacha, the main road, and the Okavango Delta; (B) Human resource selection map for firewood collection; (C) Nighttime elephant resource selection map; (D) Daytime elephant resource selection map; (E) Map of Spearman's correlation coefficients calculated between the firewood collection map and the nighttime elephant resource selection map, using a 9 × 9 moving window; (F) Map of Spearman's correlation coefficients calculated between the firewood collection map and the daytime elephant resource selection map, using a 9 × 9 moving window.
We found that during the day, all vegetation classes were significant predictors for elephant presence. Distance to settlements and the Delta were negatively selected for during the day. NDVI was positively selected for, as well as increasing distance from roads. However, at night, NDVI and distance to roads were negatively selected for, and vegetation classes became less significant predictors of elephant presence (Figure 4). Elephants also selected for closer distances to villages during the night than they did during the day.
All firewood collection occurred in the daytime. By comparing firewood collection activity to elephants' daytime and nighttime movements, we can see how the patterns could change if there was no temporal partitioning of resource use. The raster values predicted by these models were negatively correlated when comparing firewood collection and daytime elephant resource selection, and positively correlated when comparing firewood collection and nighttime elephant resource selection (Spearman's correlation coefficient: day = −0.050; night = 0.401). Spatial representation of the correlation highlights that during the daytime, human and elephant patterns of use are more negatively correlated, but that elephants' resource use at night shows more positive spatial correlation with firewood collection patterns (Figure 4).
Discussion
Interactions between people and wildlife framed only through the lens of conflict may fail to account for instances where there are neutral or beneficial interactions. Much of the research currently available on human-elephant interactions in southern Africa focuses on conflict-mitigation, in particular around farms (O'Connell-Rodwell et al., 2000; Jackson et al., 2008; Gupta, 2013), or benefits of elephants in community-based natural resource management programs (CBNRM), in particular those that rely on wildlife tourism (Turner, 2004; Mbaiwa and Stronza, 2010; Garner, 2012; Pienaar et al., 2013; Mbaiwa, 2015). Instead, our study focused on the human-elephant interactions through use of trees as a common pool resource.
People benefitted from a renewable supply of firewood created by elephants. Potentially dangerous interactions were self-mitigated through differing patterns of spatial and temporal use of the tree resources. We found temporal partitioning facilitated a situation where people could benefit from the availability of firewood generated from elephant-damaged trees by collecting wood when elephants were unlikely to be present. The tree branches that people most frequently collected for firewood were of the same tree species that elephants often damaged. People also collected firewood in the same areas where elephants moved, and some areas of the landscape had high likelihood of selection both by people for firewood and by elephants for foraging. There was a positive correlation between the areas that elephants selected at night as areas people used for firewood collection during the day. By utilizing similar areas at different times, people and elephants likely reduced their direct interactions and potential for conflict.
We found that elephants significantly impacted the vegetation in this area, damaging many trees across the study site. A majority of tree species were damaged with high frequency. When elephants browse or rub on trees, they can break branches and even trunks, and previous research has emphasized how this vegetation damage can change ecosystem structure (Mosugelo et al., 2002; Skarpe et al., 2004; de Beer et al., 2006). We connected the impact of elephant browsing to the human system by explicitly showing that elephants damage the species that people collect, therefore facilitating human livelihoods through generation of readily available firewood.
In this region we found that elephants and humans used the same tree species, which could be driven by a variety of factors. One potential cause is the abundance of wood generated from elephant damage to certain species, and the resulting availability of those species for collection. Dichrostachys cinerea, Combretum collinum, and Terminalia sericea were species frequently damaged by elephants, frequently collected for firewood, and which people expressed preference for during focal follows. Future research is necessary to understand how factors other than availability drive overlapping tree resource use by people and elephants. For example, there were some species which elephants damaged which were not collected by people. Lonchocarpus capassa is one tree species that was frequently damaged by elephants and therefore widely available for harvest, but was mostly left uncollected due to strong BaHambukushu and BaYeyi cultural taboos, in particular a belief that burning L. capassa will result in a household's livestock infertility or death.
We found little overlap in the temporal patterns of people and elephants, which is likely due to mutual avoidance. Our findings support the existing literature on elephant movement outside of protected areas (Douglas-Hamilton et al., 2005; Graham et al., 2009) but are among the first to directly include human movement in the same analysis. Elephants moved near to the settlements most frequently during dusk, night, and early morning. People collected firewood most often at mid-morning and mid-afternoon, when elephants were unlikely to be present near the village. This temporal pattern for firewood collection may be due to elephant-related safety concerns, combined with balancing timing and effort of other daily activities.
We also interpreted elephants' nocturnality around the settlements to indicate that elephants modified their behavior to avoid people and the associated risk. Although we expected to find elephants positively selecting for farther distances from settlements during the day, in fact we only found a reduced negative selection based on our model. This indicates that there are potential reasons elephants would not show high avoidance of settlements during the day. There are agricultural fields around many settlements in the region, and crops are raided by elephants. Additionally, most settlements in the Panhandle, including Mokgacha, are close to the Delta. We would expect elephants to pass near the settlements on their way to drink water and for thermoregulation throughout the day, and perhaps slightly more during the hottest part of day. We did find that wetland habitat and distance to the Delta were stronger predictors of elephant presence in the day than at night. This means elephants were selecting areas close to the Delta and wetland habitat. However, according to the patterns of temporal proximity separate from our model, elephants were rarely found near the settlements during hot midday or afternoon. Further research would be required to see if elephants exhibited this sort of pattern near water in areas where settlements are less dense in order to test whether elephants are modifying their behavior specifically due to settlements. Additionally, accounting for seasonal differences in temperature and distribution of available water may play a role in determining elephant proximity to settlements near the Delta.
Further support for temporal partitioning comes from the positive correlation between areas with high probability for firewood collection and areas where elephants were likely to be present. When elephants moved through the natural areas from dusk until dawn, browsing and damaging trees, they created an abundance of downed branches. During the day, people moved through many of the same areas where elephants moved at night. Due to elephant activity in those areas, people were able to meet their firewood harvest demands by picking up downed branches. The spatial patterns of where elephants went during the day was slightly negatively correlated with areas of human firewood collection. At night, elephants select positively for proximity to roads, settlements, and negatively for NDVI, leading to the positive correlation with firewood collection areas. If elephants exhibited the same patterns of movement during the day as they do at night, there would likely be more direct encounters between people and elephants as they would be more spatially proximate to each other.
While we recognize the often-enormous costs for people who share the environment with elephants, our study shows that human-elephant interactions around common pool resources may simultaneously carry benefits for rural, natural-resource dependent communities. Further studies that explore other overlooked aspects of human-elephant interactions can provide more evidence of complex interactions, and may one day be used to build a more complete typology that captures the kinds of interactions around resources with different governance arrangements. As humans and wildlife increasingly come into contact within social-ecological systems, it is important to implement an interdisciplinary approach to support coexistence.
Ethics Statement
This study was carried out in accordance with the recommendation of the Animal Use Protocol, Texas A&M AgriLife Research Agricultural Animal Care and Use Committee. The protocol was approved by the Agricultural Animal Care and Use Committee, AUP #2014-005A and 2017-010A. Proper veterinary and immobilization permits and procedures were approved by the Government of Botswana Department of Wildlife and National Parks, [EWT8/36/4 XVII(79)]. This study was also approved by the Texas A&M Institutional Review Board for ethnographic research including focal follows under study number #IRB2016-0255D. Verbal consent procedure and assessment of risks were approved by the board.
Author Contributions
EB and LR conceived of the research, conducted fieldwork, and co-authored the manuscript. EB analyzed the data. AmS, GM, AnS, and LF provided supervision, advice, and editing of the manuscript. All authors discussed results and contributed to final manuscript.
Funding
National Science Foundation-NSF-DGE-0654377, Applied Biodiversity Science Program for Texas A&M, provided Collaborative Multidisciplinary Research Award for research costs for EB and LR. Fulbright US Research Grant-provided funding for LR to live and conduct research in Botswana. Howard G. Buffett Foundation-provided funding for the Ecoexist Project and collars. The open access publishing fees for this article have been covered by the Texas A&M University Open Access to Knowledge Fund (OAKFund), supported by the University Libraries and the Office of the Vice President for Research.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research would not have been possible without the support of the leaders and residents of Mokgacha Village and Danga, Tinxo, Mawana, Nxininha, and Kavumo cattleposts. The authors would like to acknowledge the contributions of Ipolokeng Katholo, Susanne Vogel, Olorato Ratama, the Okavango Research Institute, Ecoexist Project, and Lethatha Gaborekwe. This study has approval from the Texas A&M Institutional Review Board, study #IRB2016- 0255D.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00117/full#supplementary-material
References
Agarwal, B. (1986). Cold Hearths and Barren Slopes: The Woodfuel Crisis in the Third World. London: Zed Books.
Alvard, M. S. (1993). Testing the “ecologically noble savage” hypothesis: interspecific prey choice by Piro hunters of Amazonian Peru. Hum. Ecol. 21, 355–387. doi: 10.1007/BF00891140
Arnold, M., Köhlin, G., and Persson, R. (2006). Woodfuels, livelihoods, and policy interventions: changing perspectives. World Dev. 34, 596–611. doi: 10.1016/J.WORLDDEV.2005.08.008
Arnold, M., Köhlin, G., Persson, R., and Shepherd, G. (2003). Fuelwood Revisited: What Has Changed in the Last Decade? Available online at: www.cifor.cgiar.org/publications/papers (accessed February 13, 2019).
Bates, D., Mächler, M., Zurich, E., Bolker, B. M., and Walker, S. C. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48 doi: 10.18637/jss.v067.i01
Ben-Shahar, R. (1993). Patterns of elephant damage to vegetation in northern Botswana. Biol. Conserv. 65, 249–256. doi: 10.1016/0006-3207(93)90057-8
Botswana Central Statistics Office (2011). 2011 Population and Housing Census. Gabarone: Botswana Central Statistics Office.
Bouget, C., Lassauce, A., and Jonsell, M. (2012). Effects of fuelwood harvesting on biodiversity — a review focused on the situation in Europe. Can. J. For. Res. 42, 1421–1432. doi: 10.1139/x2012-078
Boundja, R. P., and Midgley, J. J. (2010). Patterns of elephant impact on woody plants in the Hluhluwe-Imfolozi park, Kwazulu-Natal, South Africa. Afr. J. Ecol. 48, 206–214. doi: 10.1111/j.1365-2028.2009.01104.x
Boyce, M. S., Vernier, P. R., Nielsen, S. E., and Schmiegelow, F. K. A. (2002). Evaluating resource selection functions. Ecol. Modell. 157, 281–300. doi: 10.1016/S0304-3800(02)00200-4
Brais, S., and Drouin, P. (2012). Interactions between deadwood and soil characteristics in a natural boreal trembling aspen – jack pine stand. Can. J. For. Res. 42, 1456–1466. doi: 10.1139/x2012-061
Calenge, C., Maillard, D., Gaillard, J.-M., Merlot, L., and Peltier, R. (2002). Elephant damage to trees of wooded savanna in Zakouma National Park, Chad. J. Trop. Ecol. 18, 599–614. doi: 10.1017/S0266467402002390
Ceauşu, S., Graves, R. A., Killion, A. K., Svenning, J.-C., and Carter, N. H. (2019). Governing trade-offs in ecosystem services and disservices to achieve human-wildlife coexistence. Conserv. Biol. doi: 10.1111/cobi.13241. [Epub ahead of print].
Cerling, T. E., Wittemyer, G., Rasmussen, H. B., Vollrath, F., Cerling, C. E., Robinson, T. J., et al. (2006). Stable isotopes in elephant hair document migration patterns and diet changes. Proc. Natl. Acad. Sci. U. S. A. 103, 371–373. doi: 10.1073/pnas.0509606102
Cline-Cole, R. (2007). Woodfuel Discourses and the Re-framing of Wood Energy. Forum Dev. Stud. 34, 121–153. doi: 10.1080/08039410.2007.9666368
Cornwell, W. K., Cornelissen, J. H. C., Allison, S. D., Bauhus, J., Eggleton, P., Preston, C. M., et al. (2009). Plant traits and wood fates across the globe: rotted, burned, or consumed? Glob. Chang. Biol. 15, 2431–2449. doi: 10.1111/j.1365-2486.2009.01916.x
de Beer, Y., Kilian, W., Versfeld, W., and van Aarde, R. J. (2006). Elephants and low rainfall alter woody vegetation in Etosha National Park, Namibia. J. Arid Environ. 64, 412–421. doi: 10.1016/j.jaridenv.2005.06.015
DeMotts, R., and Hoon, P. (2012). Whose Elephants? Conserving, compensating, and competing in Northern Botswana. Soc. Nat. Resour. 25, 837–851. doi: 10.1080/08941920.2011.638362
Dewees, P. A. (1989). The woodfuel crisis reconsidered: observations on the dynamics of abundance and scarcity. World Dev. 17, 1159–1172. doi: 10.1016/0305-750X(89)90231-3
Didan, K. (2015). MODIS Vegetation Index Products MOD13Q1. Available online at: https://modis.gsfc.nasa.gov/data/dataprod/mod13.php (accessed September 19, 2018).
Douglas-Hamilton, I., Krink, T., and Vollrath, F. (2005). Movements and corridors of African elephants in relation to protected areas. Naturwissenschaften 92, 158–163. doi: 10.1007/s00114-004-0606-9
Dovie, D. B. K., Witkowski, E. T. F., and Shackleton, C. M. (2004). The fuelwood crisis in southern africa – relating fuelwood use to livelihoods in a rural village. GeoJournal 60, 123–133. doi: 10.1023/B:GEJO.0000033597.34013.9f
Dunham, K. M., Ghiurghi, A., Cumbi, R., and Urbano, F. (2010). Human–wildlife conflict in Mozambique: a national perspective, with emphasis on wildlife attacks on humans. Oryx 44, 185–193. doi: 10.1017/S003060530999086X
Eckholm, E. (1975). The Other Energy Crisis: Firewood. Worldwatch paper no. 1. Washington, DC: Worldwatch Inst.
Evans, J., and Ram, K. (2018). spatialEco: Spatial Analysis and Modelling Utilities. R package version 1.1-1.
Frank, B. (2016). Human–wildlife conflicts and the need to include tolerance and coexistence: an introductory comment. Soc. Nat. Resour. 29, 738–743. doi: 10.1080/08941920.2015.1103388
Fritz, H., Duncan, P., Gordon, I. J., and Illius, A. W. (2002). Megaherbivores influence trophic guilds structure in African ungulate communities. Oecologia 131, 620–625. doi: 10.1007/s00442-002-0919-3
Garner, K.-A. (2012). CBNRM in Botswana: The Failure of CBNRM for the Indigenous San, the Village of Xai Xai and the Wildlife of Botswana. Available online at: http://atrium.lib.uoguelph.ca/xmlui/handle/10214/4053?show=full (accessed February 5, 2019).
Goren-Inbar, N., Feibel, C. S., Verosub, K. L., Melamed, Y., Kislev, M. E., Tchernov, E., et al. (2000). Pleistocene milestones on the out-of-africa corridor at gesher benot Ya'aqov, Israel. Science 289, 944–947. doi: 10.1126/science.289.5481.944
Graham, M. D., Douglas-Hamilton, I., Adams, W. M., and Lee, P. C. (2009). The movement of African elephants in a human-dominated land-use mosaic. Anim. Conserv. 12, 445–455. doi: 10.1111/j.1469-1795.2009.00272.x
Graham, M. D., Notter, B., Adams, W. M., Lee, P. C., and Ochieng, T. N. (2010). Patterns of crop-raiding by elephants, Loxodonta africana, in Laikipia, Kenya, and the management of human-elephant conflict. Syst. Biodivers. 8, 435–445. doi: 10.1080/14772000.2010.533716
Guldemond, R., and Van Aarde, R. (2008). A meta-analysis of the impact of african elephants on savanna vegetation. J. Wildl. Manage. 72, 892–899. doi: 10.2193/2007-072
Gumbricht, T., McCarthy, J., and McCarthy, T. S. (2004). Channels, wetlands and islands in the Okavango Delta, Botswana, and their relation to hydrological and sedimentological processes. Earth Surf. Process. Landforms 29, 15–29. doi: 10.1002/esp.1008
Gupta, A. C. (2013). Elephants, safety nets and agrarian culture: understanding human-wildlife conflict and rural livelihoods around Chobe National Park, Botswana. J. Polit. Ecol. 20, 238–250. doi: 10.2458/v20i1.21766
Haynes, G. (2012). Elephants (and extinct relatives) as earth-movers and ecosystem engineers. Geomorphology 157–158, 99–107. doi: 10.1016/J.GEOMORPH.2011.04.045
Hiemstra-van der Horst, G., and Hovorka, A. J. (2009). Fuelwood: the “other” renewable energy source for Africa? Biomass Bioenergy 33, 1605–1616. doi: 10.1016/J.BIOMBIOE.2009.08.007
Ihwagi, F. W., Vollrath, F., Chira, R. M., Douglas-Hamilton, I., and Kironchi, G. (2010). The impact of elephants, Loxodonta africana, on woody vegetation through selective debarking in Samburu and Buffalo Springs National Reserves, Kenya. Afr. J. Ecol. 48, 87–95. doi: 10.1111/j.1365-2028.2009.01089.x
Jachmann, H., and Croes, T. (1991). Effects of browsing by elephants on the combretum/terminalia woodland at the nazinga game ranch, Burkina Faso, West Africa. Biol. Conserv. 57, 13–24.
Jachowski, D. S., Montgomery, R. A., Slotow, R., and Millspaugh, J. J. (2013). Unravelling complex associations between physiological state and movement of African elephants. Funct. Ecol. 27, 1166–1175. doi: 10.1111/1365-2435.12118
Jackson, T. P., Mosojane, S., Ferreira, S. M., and van Aarde, R. J. (2008). Solutions for elephant Loxodonta africana crop raiding in northern Botswana: moving away from symptomatic approaches. Oryx 42, 83–91. doi: 10.1017/S0030605308001117
Jacobs, J. M., and Work, T. T. (2012). Linking deadwood-associated beetles and fungi with wood decomposition rates in managed black spruce forests. Can. J. For. Res. 42, 1477–1490. doi: 10.1139/x2012-075
Jadhav, S., and Barua, M. (2012). The elephant vanishes: impact of human–elephant conflict on people's wellbeing. Health Place 18, 1356–1365. doi: 10.1016/j.healthplace.2012.06.019
Jones, C. G., Lawton, J. H., and Shachak, M. (1994). Organisms as ecosystem engineers. Oikos 69:373. doi: 10.2307/3545850
Jonsson, B. G., Kruys, N., Jonsson, T. R., Kruys, B. G., and Ranius, N. (2005). Ecology of species living on dead wood - lessons for dead wood management. Silva Fenn. 39, 289–309. doi: 10.14214/sf.390
Joseph, G. S., Cumming, G. S., Cumming, D. H. M., Mahlangu, Z., Altwegg, R., and Seymour, C. L. (2011). Large termitaria act as refugia for tall trees, deadwood and cavity-using birds in a miombo woodland. Landsc. Ecol. 26, 439–448. doi: 10.1007/s10980-011-9572-8
Kansky, R., Kidd, M., and Knight, A. T. (2016). A wildlife tolerance model and case study for understanding human wildlife conflicts. Bioc 201, 137–145. doi: 10.1016/j.biocon.2016.07.002
Kgathi, D., and Mlotshwa, C. (1997). “Fuelwood procurement, consumption and substitutions in selected areas of Botswana,” in Biomass Energy Policy in Africa: Selected Case Studies, eds D. Kgathi, D. Hall, A. Hategeka, and M. Sekhwela (London: Zed Books Ltd.), 10–63.
Kinahan, A. A., Pimm, S. L., and van Aarde, R. J. (2007). Ambient temperature as a determinant of landscape use in the savanna elephant, Loxodonta africana. J. Therm. Biol. 32, 47–58. doi: 10.1016/j.jtherbio.2006.09.002
Laws, R. M. (1970). Elephants as agents of habitat and landscape change in east africa. Oikos 21:1. doi: 10.2307/3543832
Leach, G., and Mearns, R. (1988). Beyond the Woodfuel Crisis: People, land and trees in Africa. London: Earthscan.
Lewis, D. (1991). Observations of tree growth, woodland structure and elephant damage on Colophospermum mopane in Luangwa Valley, Zambia. Afr. J. Ecol. 29, 207–221. doi: 10.1111/j.1365-2028.1991.tb01003.x
Mackenzie, C. A., and Ahabyona, P. (2012). Elephants in the garden: Financial and social costs of crop raiding. Ecol. Econ. 75, 72–82. doi: 10.1016/j.ecolecon.2011.12.018
Manly, B. F. J., Davis, A. J., McDonald, L. L., and Thomas, D. L. (2002). Resource Selection by Animals: Statistical Design and Analysis for Field Studies. New York, NY: Kluwer Academic Publishers.
Mapaure, I., and Ndeinoma, A. (2011). Impacts of local-level utilization pressure on the structure of mopane woodlands in Omusati region, Northern Namibia. African J. Plant Sci. 5, 305–313. Available online at: https://academicjournals.org/journal/AJPS/article-full-text-pdf/65C6C829649
Mariki, S. B., Svarstad, H., and Benjaminsen, T.A. (2015). Elephants over the cliff: explaining wildlife killings in Tanzania. Land Use Policy. 44, 19–30. doi: 10.1016/j.landusepol.2014.10.018
Mayberry, A. L., Hovorka, A. J., and Evans, K. E. (2017). Well-being impacts of human-elephant conflict in khumaga, botswana: exploring visible and hidden dimensions. Conserv. Soc. 15:280. doi: 10.4103/CS.CS_16_132
Mbaiwa, J. E. (2015). “Community-based natural resource management in Botswana,” in Institutional Arrangements for Conservation, Development and Tourism in Eastern and Southern Africa, eds R. van der Duim, M. Lamers, and J. van Wijk (Dordrecht: Springer Netherlands), 59–80.
Mbaiwa, J. E., and Stronza, A. L. (2010). The effects of tourism development on rural livelihoods in the Okavango Delta, Botswana. J. Sustain. Tour. 18, 635–656. doi: 10.1080/09669581003653500
Mills Busa, J. H. (2013). Deforestation beyond borders: addressing the disparity between production and consumption of global resources. Conserv. Lett. 6, 192–199. doi: 10.1111/j.1755-263X.2012.00304.x
Milton, S. J., and Dean, W. R. J. (1996). Rates of wood and dung disintegration in arid South African rangelands. African J. Range Forage Sci. 13, 89–93. doi: 10.1080/10220119.1996.9647904
Mmopelwa, G., Blignaut, J., and Hassan, R. (2009). Direct use values of selected vegetation resources in the Okavango Delta Wetland. South African J. Econ. Manag. Sci. 12, 242–255. Available online at: http://www.scielo.org.za/scielo.php?script=sci_arttext&pid=S2222-34362009000200008
Mole, M. A., Rodrigues DÁraujo, S., van Aarde, R. J., Mitchell, D., and Fuller, A. (2016). Coping with heat: behavioural and physiological responses of savanna elephants in their natural habitat. Conserv. Physiol. 4:cow044. doi: 10.1093/conphys/cow044
Mosugelo, D. K., Moe, S. R., Ringrose, S., and Nellemann, C. (2002). Vegetation changes during a 36-year period in northern Chobe National Park, Botswana. Afr. J. Ecol. 40, 232–240. doi: 10.1046/j.1365-2028.2002.00361.x
Nasseri, N. A., McBrayer, L. D., and Schulte, B. A. (2011). The impact of tree modification by African elephant (Loxodonta africana) on herpetofaunal species richness in northern Tanzania. Afr. J. Ecol. 49, 133–140. doi: 10.1111/j.1365-2028.2010.01238.x
National Council on Population and Development (1997). National Population Policy. Gabarone: National Council on Population and Development.
Naughton-Treves, L. (1997). Farming the forest edge: vulnerable places and people around kibale national park, Uganda. Am. Geogr. Soc. 87, 27–46.
Obanda, V., Ndeereh, D., Mijele, D., Lekolool, I., Chege, S., Gakuya, F., et al. (2008). Injuries of free ranging African elephants (Loxodonta africana africana) in various ranges of Kenya. Pachyderm 44, 54–58
O'Connell-Rodwell, C. E., Rodwell, T., Rice, M., and Hart, L. A. (2000). Living with the modern conservation paradigm: can agricultural communities co-exist with elephants? A five-year case study in East Caprivi, Namibia. Biol. Conserv. 93, 381–391. doi: 10.1016/S0006-3207(99)00108-1
O'Connor, T. G., Goodman, P. S., and Clegg, B. (2007). A functional hypothesis of the threat of local extirpation of woody plant species by elephant in Africa. Biol. Conserv. 136, 329–345. doi: 10.1016/j.biocon.2006.12.014
Ogra, M. V. (2008). Human-wildlife conflict and gender in protected area borderlands: a case study of costs, perceptions, and vulnerabilities from Uttarakhand (Uttaranchal), India. Geoforum 39, 1408–1422. doi: 10.1016/j.geoforum.2007.12.004
Olou, B. A., Yorou, N. S., Striegel, M., Bässler, C., and Krah, F.-S. (2019). Effects of macroclimate and resource on the diversity of tropical wood-inhabiting fungi. For. Ecol. Manage. 436, 79–87. doi: 10.1016/J.FORECO.2019.01.016
Osipova, L., Okello, M. M., Njumni, S. J., Ngene, S., Western, D., Hayward, M. W., et al. (2018). Fencing solves human-wildlife conflict locally but shifts problems elsewhere: a case study using functional connectivity modelling of the african elephant. J. Appl. Evol. 55. 2673–2684. doi: 10.1111/1365-2664.13246
Owen-Smith, N., and Chafota, J. (2012). Selective feeding by a megaherbivore, the African elephant (Loxodonta africana). J. Mammal. 93, 698–705. doi: 10.1644/11-MAMM-A-350.1
Pienaar, E. F., Jarvis, L. S., and Larson, D. M. (2013). Creating direct incentives for wildlife conservation in community-based natural resource management programmes in botswana. J. Dev. Stud. 49, 315–333. doi: 10.1080/00220388.2012.720366
Pozo, R. A., Coulson, T., McCulloch, G., Stronza, A. L., and Songhurst, A. C. (2017). Determining baselines for human-elephant conflict: a matter of time. PLoS One 12, 1–17. doi: 10.1371/journal.pone.0178840
Pringle, R. M. (2008). Elephants as agents of habitat creation for small vertebrates at the patch scale. Ecology 89, 26–33. doi: 10.1890/07-0776.1
Pröpper, M., Gröngröft, A., Finckh, M., Stirn, S., Cauwer, V., De Lages, F., et al. (2015). The Future Okavango – Findings, Scenarios and Recommendations for Action. Research Project Final Synthesis Report, 2010 — 2015 (The Future Okavango Project), 190.
QGIS Development Team (2018). QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online at: http://qgis.osgeo.org
R Core Team (2018). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: https://www.R-project.org (accessed July 2, 2018).
Roever, C. L., van Aarde, R. J., and Chase, M. J. (2013). Incorporating mortality into habitat selection to identify secure and risky habitats for savannah elephants. Biol. Conserv. 164, 98–106. doi: 10.1016/j.biocon.2013.04.006
Shackleton, C. M. (1998). Annual production of harvestable deadwood in semi-arid savannas, South Africa. For. Ecol. Manage. 112, 139–144. doi: 10.1016/S0378-1127(98)00321-1
Shackleton, C. M., Shackleton, S. E., Buiten, E., and Bird, N. (2007). The importance of dry woodlands and forests in rural livelihoods and poverty alleviation in South Africa. For. Policy Econ. 9, 558–577. doi: 10.1016/J.FORPOL.2006.03.004
Shaheen, H., Azad, B., Mushtaq, A., and Ahmad Khan, R. W. (2016). Fuelwood consumption pattern and its impact on forest structure in Kashmir Himalayas. Bosque 37, 419–424. doi: 10.4067/S0717-92002016000200020
Sheil, D., and Salim, A. (2004). Forest tree persistence, elephants, and stem scars. Biotropica 36, 505–521. doi: 10.1111/j.1744-7429.2004.tb00346.x
Shrader, A. M., Bell, C., Bertolli, L., and Ward, D. (2012). Forest or the trees: at what scale do elephants make foraging decisions? Acta Oecol. 42, 3–10. doi: 10.1016/j.actao.2011.09.009
Signer, J., Fieberg, J., and Avgar, T. (2018). Animal Movement Tools (amt): R-Package for Managing Tracking Data and Conducting Habitat Selection Analyses. arXive:1–27.
Sitati, N. W., Walpole, M. J., and Leader-Williams, N. (2005). Factors affecting susceptibility of farms to crop raiding by African elephants: using a predictive model to mitigate conflict. J. Appl. Ecol. 42, 1175–82. doi: 10.1111/j.1365-2664.2005.01091.x
Sitati, N. W., Walpole, M. J., Smith, R. J., and Leader-Williams, N. (2003). Predicting spatial aspects of human and elephant conflict. J. Appl. Ecol. 4, 667–677. doi: 10.1046/j.1365-2664.2003.00828.x
Skarpe, C., Aarrestad, P. A., Andreassen, H. P., Dhillion, S. S., Dimakatso, T., du Toit, J. T., et al. (2004). The return of the giants: ecological effects of an increasing elephant population. Ambio 33, 276–282. doi: 10.2307/4315497
Songhurst, A. C. (2014). Ecoexist Report on Elephant Collaring Exercise Eastern and Western Panhandle. Botswana: Ecoexist Project.
Songhurst, A. C., McCulloch, G., and Coulson, T. (2016). Finding pathways to human-elephant coexistence: a risky business. Oryx 50, 713–720. doi: 10.1017/S0030605315000344
Stokke, S., and du Toit, J. T. (2000). Sex and size related differences in the dry season feeding patterns of elephants in Chobe National Park, Botswana. Ecography 23, 70–80. doi: 10.1111/j.1600-0587.2000.tb00262.x
Sukumar, R. (2003). The Living Elephants: Evolutionary Ecology, Behaviour, and Conservation. New York, NY: Oxford University Press.
Tietema, T. (1993). Biomass determination of fuelwood trees and bushes of Botswana, Southern Africa. For. Ecol. Manage. 60, 257–269. doi: 10.1016/0378-1127(93)90083-Y
Tietema, T., Ditlhogo, M., Tibone, C., and Mathalaza, N. (1991). Characteristics of eight firewood species of Botswana. Biomass Bioenergy 1, 41–46. doi: 10.1016/0961-9534(91)90050-M
Turner, S. (2004). “The commons in an age of global transition: challenges, risks and opportunities,” in the Tenth Biennial Conference of the International Association for the Study of Common Property. in 10th IASCP Conference (Oaxaca), 9–13.
Twine, W., and Magome, H. (2007). “Interactions between elephants and people.” in Elephant management: A scientific assessment of South Africa, eds. R. J. Scholes and K. G. Mennel (Johannesburg: Witwatersrand University Press), 206–240.
Vanak, A. T., Shannon, G., Thaker, M., Page, B., Grant, R., and Slotow, R. (2012). Biocomplexity in large tree mortality: interactions between elephant, fire and landscape in an African savanna. Ecography 35, 315–321. doi: 10.1111/j.1600-0587.2011.07213.x
Vázquez, D. P., Alvarez, J. A., Debandi, G., Aranibar, J. N., and Villagra, P. E. (2011). Ecological consequences of dead wood extraction in an arid ecosystem. Basic Appl. Ecol. 12, 722–732. doi: 10.1016/J.BAAE.2011.08.009
Keywords: human-elephant conflict, African elephants, human-wildlife interactions, social-ecological systems, Botswana, resource selection function
Citation: Buchholtz EK, Redmore L, Fitzgerald LA, Stronza A, Songhurst A and McCulloch G (2019) Temporal Partitioning and Overlapping Use of a Shared Natural Resource by People and Elephants. Front. Ecol. Evol. 7:117. doi: 10.3389/fevo.2019.00117
Received: 30 November 2018; Accepted: 22 March 2019;
Published: 11 April 2019.
Edited by:
Laura Kehoe, University of Victoria, CanadaReviewed by:
Bruce Alexander Schulte, Western Kentucky University, United StatesSamrat Mondol, Wildlife Institute of India, India
Copyright © 2019 Buchholtz, Redmore, Fitzgerald, Stronza, Songhurst and McCulloch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Erin K. Buchholtz, ZWtidWNoaG9sdHpAZ21haWwuY29t