 Mackenzie E. Davenport1,2
Mackenzie E. Davenport1,2 Michael B. Bonsall
Michael B. Bonsall Hope Klug
Hope Klug- 1Department of Biology, Geology, and Environmental Science, University of Tennessee at Chattanooga, Chattanooga, TN, United States
- 2Honors College, University of Tennessee at Chattanooga, Chattanooga, TN, United States
- 3Mathematical Ecology Research Group, Department of Zoology, University of Oxford, Oxford, United Kingdom
- 4St Peter's College, University of Oxford, Oxford, United Kingdom
- 5SimCenter, University of Tennessee at Chattanooga, Chattanooga, TN, United States
Parental care is a key life-history trait that increases offspring fitness. When one thinks of parental care, nurturing behaviors such as guarding, provisioning, and grooming typically come to mind. However, such conventional forms of care often co-occur with offspring abandonment or filial cannibalism (the consumption of one's offspring). Offspring abandonment and filial cannibalism are typically viewed as evolutionary conundrums that are contradictory to parental care. Here, we hypothesize that when offspring survival is density dependent, offspring abandonment and filial cannibalism can themselves function as forms of parental care for remaining offspring. We use a mathematical model to test this hypothesis. Our results suggest that offspring abandonment and filial cannibalism can function as forms of parental care. These results have the potential to broaden our general understanding of what is considered to be parental care.
Introduction
Parental care is a fundamental life-history trait found in many animals (Clutton-Brock, 1991; Royle et al., 2012). Parental care increases offspring fitness and includes behaviors such as egg cleaning (Knouft et al., 2003; Alonso-Alvarez and Velando, 2012; Klug and Bonsall, 2014), offspring guarding (Klug et al., 2005; Buzatto et al., 2007; Kölliker, 2007; Alonso-Alvarez and Velando, 2012; Klug and Bonsall, 2014), provisioning of young (Simmons and Parker, 1989; Kölliker, 2007; Alonso-Alvarez and Velando, 2012; Klug and Bonsall, 2014), and oviposition-site selection (Refsnider and Janzen, 2010).
Parental care commonly co-occurs with behaviors such as offspring abandonment and filial cannibalism, i.e., the consumption of one's own offspring (Lack, 1954; Rohwer, 1978; Clark and Wilson, 1981; FitzGerald, 1992; Manica, 2002; Payne et al., 2002; Klug and Bonsall, 2007; Zuckerman et al., 2014). Offspring abandonment and filial cannibalism have typically been viewed as evolutionary conundrums that are contradictory to parental care (Klug and Bonsall, 2007). However, it has been noted that offspring abandonment and filial cannibalism might occur in response to density-dependent offspring mortality (i.e., a situation in which offspring mortality increases as offspring density increases) (Lack, 1954; Clark and Wilson, 1981; Payne et al., 2002, 2004; Klug and Bonsall, 2007; Zuckerman et al., 2014). If egg survival is density dependent, such that removing some offspring from a clutch improves the survival of remaining offspring (see, e.g., Smith et al., 1987; Bjørnstad et al., 1999; Payne et al., 2002; Gunnarsson et al., 2006; Klug et al., 2006; Vallon et al., 2016), offspring abandonment and filial cannibalism might themselves be forms of parental care for remaining offspring.
To the best of our knowledge, the idea that offspring abandonment and filial cannibalism might themselves be and evolve as forms of parental care, even in the absence of other more conventional parental care behaviors, has not been explicitly considered. Below, we outline how offspring abandonment and filial cannibalism might serve as forms of parental care and then develop a mathematical model of offspring abandonment and filial cannibalism to evaluate this hypothesis.
Offspring of many species lay young in close proximity to one another. For example, eggs of fish are often spawned in close proximity to other eggs within a male's territory (Baylis, 1981). Likewise, even in the absence of care, communal egg laying is common in a range of insects, retiles, and amphibians (reviewed in Sean Doody et al., 2009). As a result, it is common for offspring, particularly eggs, to be laid in a given area. In addition, parental care behaviors such as provisioning, cleaning, and thermoregulation typically require offspring to be spatially clumped. Given this, we would often expect offspring density to increase early in the evolution of parental care. If offspring become clumped at the site of laying or oviposition—either because other forms of care require clumping or because eggs begin to be deposited in a given area (Baylis, 1981; Sean Doody et al., 2009)—we might expect offspring mortality to be influenced by within-clutch offspring density. When offspring are present at high density, there is potentially increased disease transmission (Vallon et al., 2016), decreased oxygen availability (Payne et al., 2002, 2004), or decreased food (Shepherd and Cushing, 1980; Frederiksen and Bregnballe, 2000), and density-dependent offspring mortality has been found in a range of systems (Smith et al., 1987; Bjørnstad et al., 1999; Payne et al., 2002; Gunnarsson et al., 2006; Vallon et al., 2016). If parents can reduce the density, and thereby the overall mortality, of their young through filial cannibalism or abandonment of young (i.e., the removal of young from the oviposition or laying site), cannibalism or abandonment might themselves function as parental care. Indeed, some previous research suggests that filial cannibalism can improve the survival of remaining offspring. For example, in the beaugregory damselfish (Stegastes leucostic), fathers were more likely to eat eggs under low oxygen conditions, and this is hypothesized to be an adaptive way for these males to reduce density and improve the survival of remaining offspring under low oxygen conditions (Payne et al., 2002).
Below, we use a theoretical model to test the hypothesis that offspring abandonment and filial cannibalism can function as forms of parental care. We explore the fitness benefits of offspring abandonment and filial cannibalism when egg mortality is affected by within-clutch egg density effects. Under certain circumstances, we expect that when egg mortality is affected by within-clutch density effects, offspring abandonment and filial cannibalism can be selected for due to increases in overall offspring survival; in contrast, we would expect offspring abandonment and filial cannibalism to be associated with no or relatively weak benefits when egg survival is not affected by within-clutch density effects. To isolate potential benefits of offspring abandonment and filial cannibalism, we focus only on these two strategies and assume that no other forms of post-hatching parental care occur.
Materials and Methods
Model Overview
Using an evolutionary ecology model following Klug and Bonsall (2010) (see also Klug and Bonsall, 2007, 2014; Bonsall and Klug, 2011a,b), a rare mutant that exhibits offspring abandonment or filial cannibalism is introduced into a resident population with no abandonment or cannibalism (equations 1–25 below). The resident population is assumed to be in ecological equilibrium (equations 3–4 below), and we ask whether the mutant exhibiting offspring abandonment or filial cannibalism can invade the resident population from rare. We determine if offspring abandonment and filial cannibalism can function as a form of parental care for the remaining offspring and result in positive fitness that would allow the mutant strategy to evolve.
In our model, individuals undergo egg and juvenile stages before maturing and reproducing as adults. Resident and mutant individuals have the same life-history parameters prior to accounting for costs and benefits of abandonment or cannibalism (described below). The mutant then differs from the resident in two ways: first, as mentioned above, we would expect offspring of many species to be clumped after laying or oviposition; as such, within-clutch density effects are expected to occur, and we therefore assume that mutant egg survival is affected by within-clutch egg density effects (Figure 1). Specifically, as within-clutch density effects increase (rm in Equation 5 below), egg mortality increases for mutants (equations 19–20 below). Second, resident and mutants differ as mutants exhibit offspring abandonment (equations 15, 17, and 18 below) or filial cannibalism (Equation 16 below).
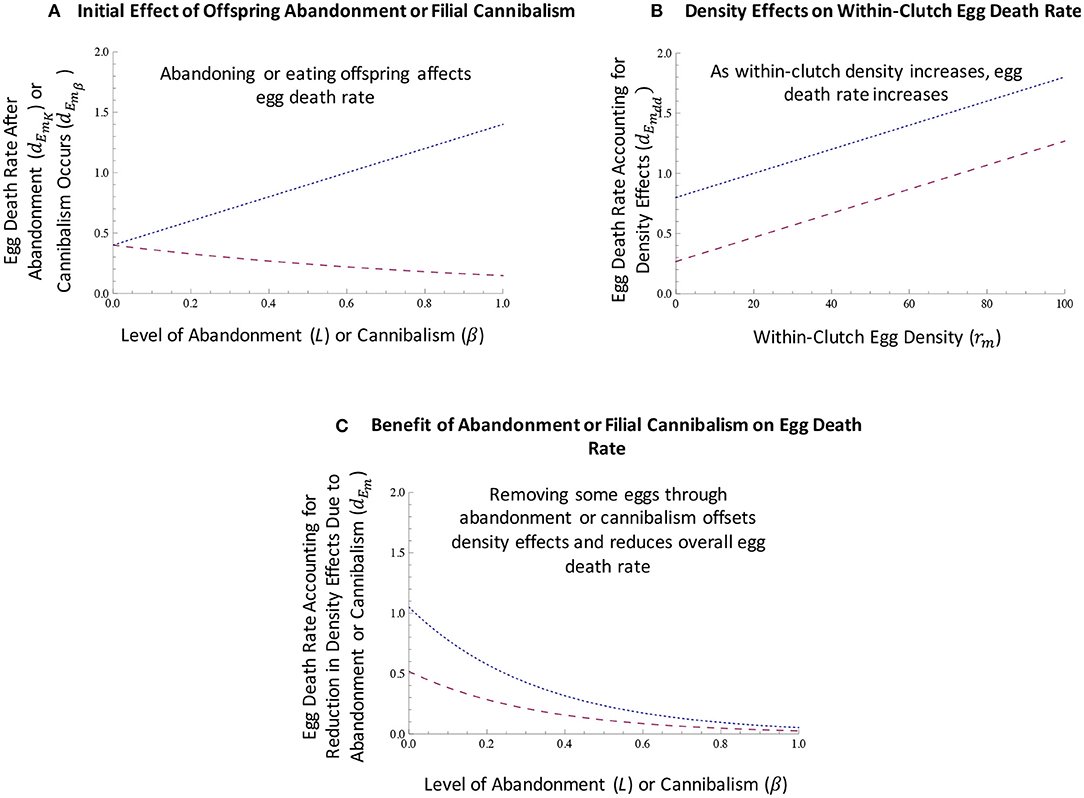
Figure 1. Egg death rate of mutants is affected by costs and benefits of abandonment and cannibalism and by density effects. (A) When parents abandon or consume some of their offspring, egg death rate might initially increase because eggs are damaged or killed when they are removed from the laying or oviposition site or as they are consumed (small dashed line; abandonment scenario 1). Alternatively, abandonment might improve egg survival (medium dashed line; abandonment scenario 2), as removed eggs could have greater survival away from the laying or oviposition site when density effects exist. Specifically, in (A) we assumed that initial, baseline egg death rate (dEm0) was equal to 0.4; as parents abandon or consume their young, the egg death rate (dEmL or dEmβ) when accounting for this abandonment (L) or cannibalism (β) increases due to the mortality caused by parents or decreases due eggs being moved to another, less dense location. (B) As offspring are assumed to be spatially clumped in the early evolution of care, we assume that there are density effects on egg death rate. Specifically, as within-clutch egg density (rm) increases, egg death rate (dEmdd) increases. In (B), we assumed that baseline egg death rate (dEm0) was equal to 0.4 and that the level of abandonment or filial cannibalism was equal to 0.4 (i.e., L = β = 0.4), which means that dEmL = dEmβ = 0.8 when abandonment and cannibalism decrease egg survival and dEmL = 0.268 when abandonment increases egg survival. In (B), we assumed that the magnitude of density effects, z, was equal to 0.01. (C) When parents abandon or consume offspring, within-clutch egg density is reduced. As a result, abandonment or filial cannibalism decrease within-clutch egg density of remaining eggs, and thereby reduce egg death rate. In (C), we again assumed that baseline egg death rate (dEm0) was 0.4, that the rate of cannibalism or abandonment (L or β) was 0.4, and that z = 0.01; we also assumed that rm = 25. The small dashed line illustrates the scenario in which cannibalism and abandonment initially increase egg death rate; the medium dashed line illustrates the scenario in which abandonment initially decreases egg death rate. See text for full details of model equations and parameters.
Filial cannibalism is always assumed to reduce offspring survival (i.e., offspring die when consumed by parents; Equation 16 below). With regard to abandonment, we consider scenarios in which being abandoned either increases or decreases offspring mortality. In the first scenario (abandonment scenario 1), abandonment increases egg mortality (Equation 15 below) but also reduces the magnitude of density effects on egg mortality when density effects exist (equations 19–20 below) (Figure 1). Such an abandonment scenario is reflective of situations in which offspring are damaged or killed when they are removed from the laying or oviposition site (i.e., eggs are damaged during the process of abandonment). Alternatively, it is possible that being abandoned could be beneficial to young when within-in clutch density effects occur. That is, when within-clutch density affects egg survival, abandoning young and removing them from a territory, nest, or given area might increase their survival. In the second abandonment scenario (abandonment scenario 2), we therefore assume that being abandoned improves survival of abandoned offspring, as they will no longer experience within-clutch density effects. In such a scenario, abandonment is assumed to increase the survival of those abandoned offspring, thereby increasing overall initial offspring survival, and we assume that overall egg survival increases as the rate of abandonment increases when density effects occur (Equation 17 below). When parents exhibit filial cannibalism, they receive energetic benefits that increase adult mutant survival (Equation 23 below); no energetic benefits are associated with offspring abandonment.
We model scenarios in which density effects either do or do not affect within-clutch egg mortality; in doing so, we determine if the density effects on egg mortality allow abandonment or cannibalism to result in positive fitness.
Our framework, which is described in more detail below, utilizes evolutionary invasion analysis (see, e.g., Ch. 12 of Otto and Day, 2007). Specifically, this approach allows us to consider whether a population that is initially fixed for a strategy (no abandonment or cannibalism) can be invaded by a mutant strategy that represents a different abandonment or cannibalism strategy. Our model and analyses therefore focus on the evolutionary origin of new strategies in a population. This allows us to gain insight into the conditions under which abandonment and filial cannibalism are most likely to result in positive fitness and be able to invade a resident population exhibiting no abandonment or cannibalism.
Full Model Dynamics
Resident Parameters
To utilize evolutionary invasion analysis, one must first identify the dynamics of the resident strategy in the absence of the mutant strategy (Otto and Day, 2007). In our model, individuals pass through an egg (E), juvenile, and adult stage (A). In the population, eggs increase as adults reproduce and decrease as eggs mature and as they die, such that
Here, r represents the rate of egg fertilization by individual adults in the population; as such, in this model, r can be thought of as a measure of within-clutch egg number. The term dE represents egg death rate and mE represents egg maturation rate. The population has a carrying capacity represented by K, and adult reproduction is restricted by density-dependence. Adults in the population increase as eggs mature and pass through the juvenile stage, and decrease as adults die, such that
Here, τ represents the length of the juvenile stage and σJ represents the rate of survival through the juvenile stage.
The next step of evolutionary invasion analysis is to determine the equilibria of the resident model (Otto and Day, 2007). The ecological equilibrium densities in the resident population will occur when In our model, the ecological equilibria are thus:
and
Mutant Dynamics and Fitness
The third step of evolutionary invasion analysis is to define the dynamics of a mutant strategy and identify the interaction between the mutant and resident individuals (Otto and Day, 2007). The following equations provide the invasion dynamics of the rare mutant:
Here, the subscript m is used to show that these variables relate to the mutant strategy of abandoning or cannibalizing offspring, and A* represents the equilibrial abundance of the resident adult population (i.e., the abundance when ). As with the resident strategy, mutant eggs increase in density as adults reproduce and decrease as eggs die or mature and leave the egg stage (Equation 5); likewise, mutant adults increase in density as mutant eggs mature, survive, and pass through the juvenile stage and decrease in density as adults die (Equation 6). As the mutant is assumed to be rare, mutant reproduction is limited by competition with adult residents (Equation 5); through this competition for resources that limit reproduction, mutant and resident individuals are assumed to interact. For mutants, parents are assumed to be spatially associated with their young (discussed above). Given this, rm can be thought of as a measure of within-clutch density for mutants.
Unless otherwise noted, all resident parameters are equal to their corresponding baseline values, which are provided below, such that:
Additionally, unless otherwise noted below, all mutant parameters are equal to their corresponding baseline values, which are provided below, such that:
In other words, except for where noted below, we assume no trade-offs on mutant or resident parameters.
Mutant Trade-Offs Associated With Abandonment and Cannibalism
When mutant parents exhibit offspring abandonment or filial cannibalism that decreases the survival of those offspring that are abandoned (abandonment scenario 1 above) or consumed, the baseline mutant egg death rate is equal to the baseline resident egg death rate plus the level of abandonment or cannibalism exhibited, such that:
where L is equal to the rate of egg abandonment, and
where β represents the rate of filial cannibalism. Thus, the rate of egg mortality initially increases as eggs are directly lost due to abandonment or cannibalism (Figure 1A).
For the scenario in which abandonment is beneficial to young (i.e., when abandonment increases survival of offspring that are abandoned, thereby improving overall offspring survival; abandonment scenario 2 above, Figure 1A), the baseline mutant egg death rate decreases as the level of abandonment increases when density effects are present, such that:
In the scenario in which we assume no negative consequence of being removed from the site of laying or oviposition (abandonment scenario 2), egg survival is assumed to be unaffected by abandonment when density effects are not present, such that:
In scenarios in which there are density effects on within-clutch egg mortality, the mutant egg death rate then increases as mutant within-clutch egg density effects (rm) increase:
for the case of abandonment, and:
for the case of filial cannibalism (Figure 1B). Here, z represents the magnitude of density effects acting on egg mortality, and the magnitude of density effects on egg mortality increases as the value of z increases (Figure 2). In the above equations 19 and 20, the only difference is whether abandonment or filial cannibalism occurs (i.e., density effects are assumed to be the same for abandonment and cannibalism).
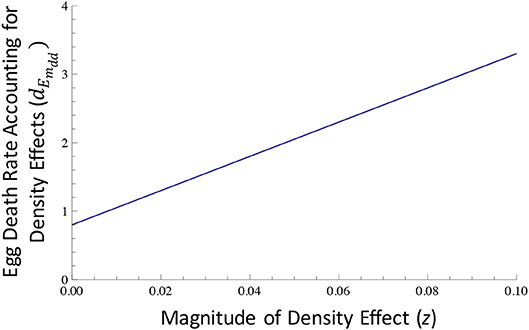
Figure 2. The magnitude of density effects in relation to egg death rate. Egg death rate of mutant individuals increases as within-clutch density effects increase (see Equations 19–20 in the text for additional details). The term z is a measure of the magnitude of these density effects. As z increases, density effects increase, and egg mortality increases for a given within-clutch egg density, rm, with increasing z. Here, we assumed that: (i) within-clutch egg density, rm, was 25; (ii) baseline egg death rate (dEm0) was equal to 0.4; and (iii) the level of abandonment or filial cannibalism was equal to 0.4 (i.e., L = β = 0.4). In this case, we assume that both cannibalism and abandonment decrease egg survival, and as a result, the egg death rate after accounting for egg loss due to abandonment or cannibalism was equal to 0.8. If we assume that z = 0.01, which is the value of z assumed in most of our analyses below, density effects would cause egg mortality rate (dEmdd) to increase to approximately 1.05. If z were larger, density effects would cause egg mortality rate to be even greater.
To determine whether density effects allow offspring abandonment and filial cannibalism to function as forms of care, we compare the fitness of abandonment and cannibalism for the scenario in which (1) density effects exist (see also Figure 1B) and (2) density effects are absent and there are no density effects on egg mortality (i.e., dEm = dEm0, as in Equation 11 above). In comparing the scenarios in which density effects are present vs. absent in mutants, we are able to determine if the density effects themselves lead to relatively high fitness of abandonment and cannibalism.
When density effects on within-clutch egg mortality exist, decreasing density via abandonment or cannibalism is assumed to offset the overall density effects on egg mortality. Specifically, the rate of abandonment or cannibalism offsets the effects of density on egg mortality (Figure 1C), such that:
for the case of offspring abandonment, and:
for the case of filial cannibalism. In equations 21 and 22, which only differ with respect to whether abandonment or cannibalism is occurring, s reflects the magnitude of the benefit of reducing egg density in relation to egg survival; when s is large, egg death rate decreases at a greater rate as eggs are removed from the nest through abandonment or cannibalism in comparison to the case in which s is relatively small.
In summary, mutant eggs experience three main effects related to mortality: (1) offspring abandonment and filial cannibalism can initially affect egg mortality as eggs are removed from the nest or consumed (Figure 1A); (2) as eggs are assumed to be spatially clumped, density effects associated with rm can also increase egg mortality (Figure 1B); and (3) removing some eggs through abandonment or cannibalism can reduce density and offset density effects, thereby reducing mortality of remaining eggs and the overall egg mortality rate (Figure 1C).
When parents abandon their offspring, we assume that offspring are killed or simply removed from the vicinity of the parent and other offspring (e.g., they might be pushed out of a nest of removed from a territory). When parents exhibit filial cannibalism, we assume that there is an energetic benefit of eating eggs that leads to increased parental survival, such that:
Here, γ represents the relative level of energetic benefits provided to the parent through filial cannibalism. In this model, γ = 0.5 is used to represent relatively low energetic benefits of cannibalism, and values of 1 and 1.5 represent moderate and high energetic benefits of cannibalism (Figure 3).
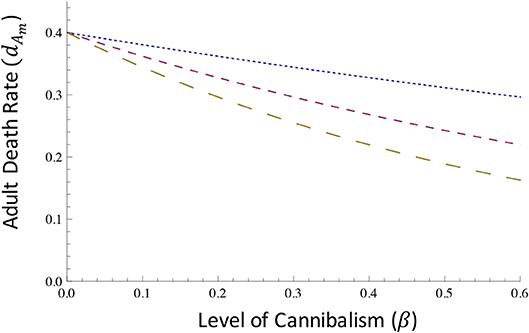
Figure 3. Energetic benefits of filial cannibalism to mutant parents. Mutant parents that exhibit filial cannibalism are assumed to gain energetic benefits that reduce their mortality (dAm). Here, we consider three levels of energetic benefits of filial cannibalism (γ): low energetic benefits to parents (small dashed line), such that dAm = dAm0*Exp[−0.5*β]; medium energetic benefits to parents (medium dashed line), such that dAm = dAm0*Exp[−1*β]; and high energetic benefits to parents (large dashed line), such that dAm = dAm0*Exp[−1.5*β]. In all cases illustrated here, we assume that dAm0 = 0.4.
Invasion Matrix and Fitness Calculation
The fourth step of evolutionary invasion analysis is to perform a local stability analysis to determine the dominant eigenvalue of the invasion matrix. The dominant eigenvalue can be thought of as the reproductive factor or growth rate of the mutant when the mutant is rare and can provide a measure of fitness of the mutant strategy (Otto and Day, 2007). In other words, the relative fitness of the mutant (i.e., the fitness of the mutant strategy relative to that of the resident) is measured as the population-level growth rate of that mutant strategy, which is the dominant eigenvalue from following matrix:
The components of the above matrix are derived from the dynamics of the linearized mutant strategy around the trivial steady state and are specifically determined by the partial derivative of the demographic dynamics of the mutant (equations 5–6 above). The dominant eigenvalue of the above matrix can then be determined by solving the resulting characteristic equation for λ. Specifically, as τ → 0, taking the determinant of expression 24 above yields the following characteristic equation:
Here, solving for λ and determining the dominant eigenvalue provides us with a Fisherian measure of the fitness of the mutant strategy relative to the resident strategy (i.e., a measure of the relative growth rate of the mutant strategy or a measure of the reproductive factor of the mutant strategy; Otto and Day, 2007). Specifically, the eigenvalues of expression 24 are the values of λ that satisfy Equation 25 above. When λ is greater than zero (that is, local stability around the trivial steady state for the mutant strategy is unstable), the mutant strategy (offspring abandonment or filial cannibalism) is associated with positive fitness and would be expected to be able to invade the resident strategy of no abandonment/no cannibalism. When fitness is negative, abandonment and cannibalism are not expected to be able to evolve.
Given that the aim of our model is to provide a first, proof-of-concept consideration of whether offspring abandonment and/or filial cannibalism can potentially increase remaining offspring survival and can therefore ever be considered a form of parental care, we considered the following fixed parameters for our analyses unless otherwise noted below:
dE0 = dEm0 = 0.4, dA0 = dAm0 = 0.4, mE = mEm = 0.4, K = Km = 1000, r = rm = 25, σJ = σJm = 0.1, τ = τm = 0.1, s = 3, and z = 0.01. It is important to note that as we use a continuous time model, mortality and maturation are quantified as rates (e.g., egg death rate is the instantaneous rate at which eggs die at time t). Thus, mortality and maturation rates do not need to be less than one to be biologically realistic. To assess the generality of our findings, we then also examined how the fitness of offspring abandonment and filial cannibalism change in relation to within-clutch egg density, the magnitude of benefit of abandoning or eating eggs in relation to within-clutch density effects, and baseline egg and adult mortality.
After determining the parameters under which offspring abandonment and filial cannibalism can potentially function as forms of parental care, it would be possible to then determine if there is a trait value that can resist invasion by other rare mutant strategies (i.e., the evolutionary stable strategy, ESS). However, the aim of our model is not to identify any particular ESS conditions, particularly since any ESS is likely to depend on the specific parameters and trade-offs associated with abandonment or cannibalism; rather, our aim is to determine if offspring abandonment and/or filial cannibalism can result in positive fitness when density effects occur and therefore function as forms of parental care.
All analyses were performed in Wolfram Mathematica version 8.0 or higher, and the Mathematica code for the model is publicly available in the Open Science Framework at the following link: https://osf.io/y4a82/.
Results
When mortality increases as within-clutch density effects increase, offspring abandonment that either increases or decreases the survival of abandoned offspring can function as a form of parental care, and fitness is positive over a range of levels of offspring abandonment (Figures 4A,B). When egg mortality does not depend on within-clutch density effects, the fitness of offspring abandonment is negative relative to the no-abandonment scenario across the parameters considered (Figure 4A). Thus, the positive fitness that results from offspring abandonment in this scenario is due to the presence of density effects on egg mortality. Offspring abandonment is particularly likely to result in fitness benefits and be selected for when the process of abandoning young improves the survival of those young that are removed from the nest, territory, and/or laying or oviposition site (Figure 4B).
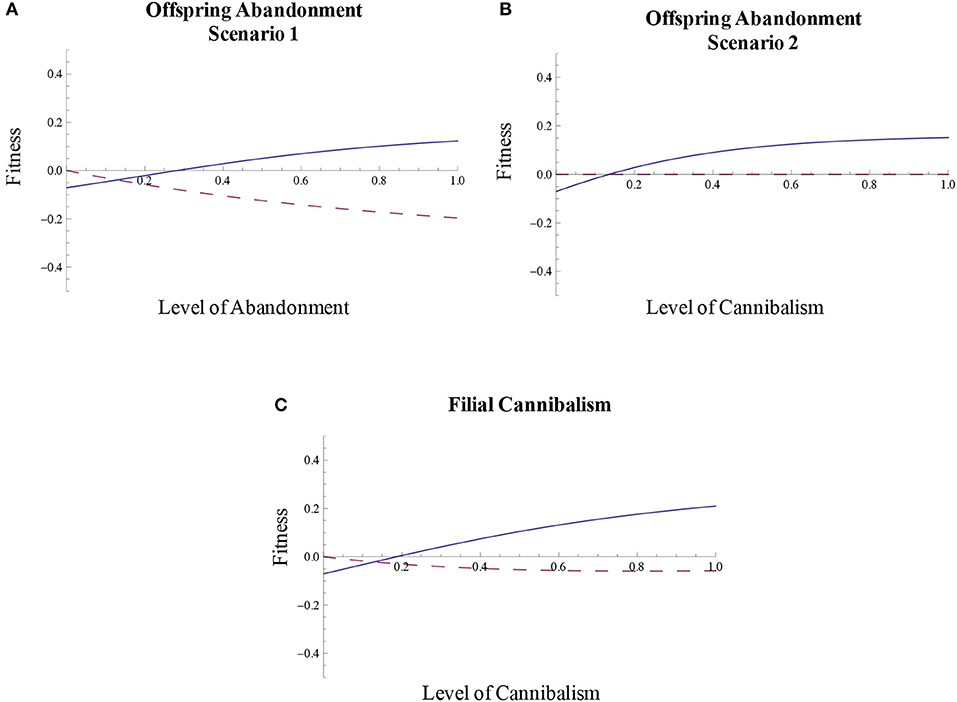
Figure 4. Offspring abandonment and filial cannibalism function as forms of parental care. When (A) egg mortality increases as within-clutch density effects increase, offspring abandonment that initially decreases egg survival (abandonment scenario 1), beyond a threshold level, is associated with positive fitness and can function as a form of parental care in some cases (solid line). If egg mortality does not increase as within-clutch density effects increase, offspring abandonment will not function as a form of parental care (dashed line). When (B) egg mortality increases as within-clutch density effects increase, offspring abandonment that initially increases egg survival (abandonment scenario 2), beyond a threshold level, is associated with positive fitness and can function as a form of parental care in some cases (solid line). If egg mortality does not increase as within-clutch density effects increase, offspring abandonment will not function as a form of parental care (dashed line, which overlaps with the x-axis). When (C) filial cannibalism reduces the density effects on egg mortality and provides energetic benefits to parents, beyond a threshold, cannibalism can be associated with positive fitness and function as a form of parental care (solid line). If filial cannibalism does not reduce density effects on egg mortality, filial cannibalism will not function as a form of care (dashed line).
With associated energetic benefits and reductions in density effects on egg mortality, filial cannibalism can also function as a form of parental care (Figure 4C). This strategy is associated with positive fitness when density effects occur compared to the scenario where there are no effects of density on egg mortality (Figure 4C). As such, the density effects on egg mortality allow filial cannibalism to function as a form of parental care. In addition, the level of energetic benefits from cannibalism also affect the fitness of cannibalism. In Figure 4C, we assume relatively low energetic benefits. If, however, the energetic benefits of cannibalism are moderate or high, filial cannibalism can be associated with positive fitness even when there are no density effects on within-clutch offspring mortality (Figures 5A,B). These results suggest that filial cannibalism can evolve as a form of parental care and/or be favored by energetic benefits.
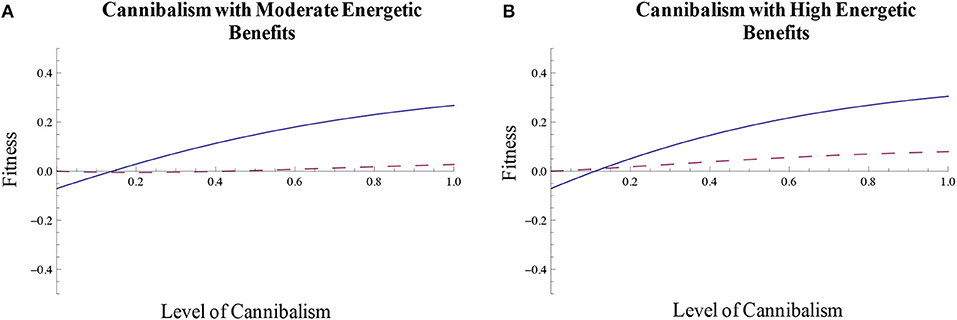
Figure 5. Energetic benefits affect the fitness of filial cannibalism. When filial cannibalism provides (A) moderate or (B) high energetic benefits to parents, fitness associated with filial cannibalism can be positive even when there are no effects of density on within-clutch mortality (dashed line). The solid line reflects the scenario in which there are density effects on egg mortality.
As one would expect, the fitness associated with offspring abandonment and filial cannibalism will be greater when the magnitude of the benefit of abandonment and cannibalism in relation to offsetting density effects is relatively large (Figures 6A,B). Likewise, the fitness associated with offspring abandonment and filial cannibalism that offset density effects on egg mortality will be greater at higher within-clutch mutant egg densities (Figures 7A,B). Offspring abandonment and filial cannibalism that offset density effects are also expected to be associated with relatively high fitness when baseline egg mortality is relatively high (Figures 7C,D). This pattern likely occurs because the offspring need for the benefits of abandonment and cannibalism will be greater at higher egg mortalities. In other words, when egg mortality is already low, there will be relatively little need or benefit of any form of parental care. Finally, the fitness benefit of offspring abandonment and filial cannibalism increases as adult death rate increases, particularly for the case of filial cannibalism (Figures 7E,F).
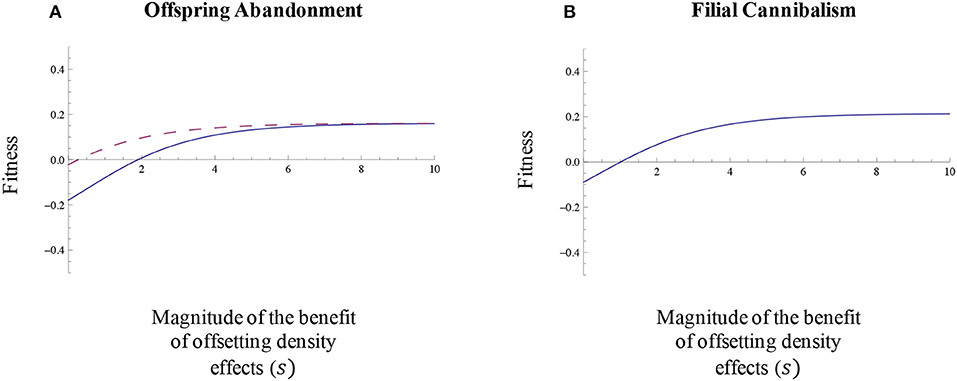
Figure 6. Magnitude of the benefit of abandonment or cannibalism in relation to offsetting density effects. The fitness associated with (A) offspring abandonment that initially decreases (abandonment scenario 1, solid line) or increases (abandonment scenario 2, dashed line) egg survival and (B) filial cannibalism that initially decreases egg survival will be greater when the magnitude of the benefit of abandonment or cannibalism in relation to offsetting density effects (s) is relatively large (see also Equations 21 and 22 in the main text for additional details). In these scenarios, β = 0.6 and all other parameter values are as described in the text.
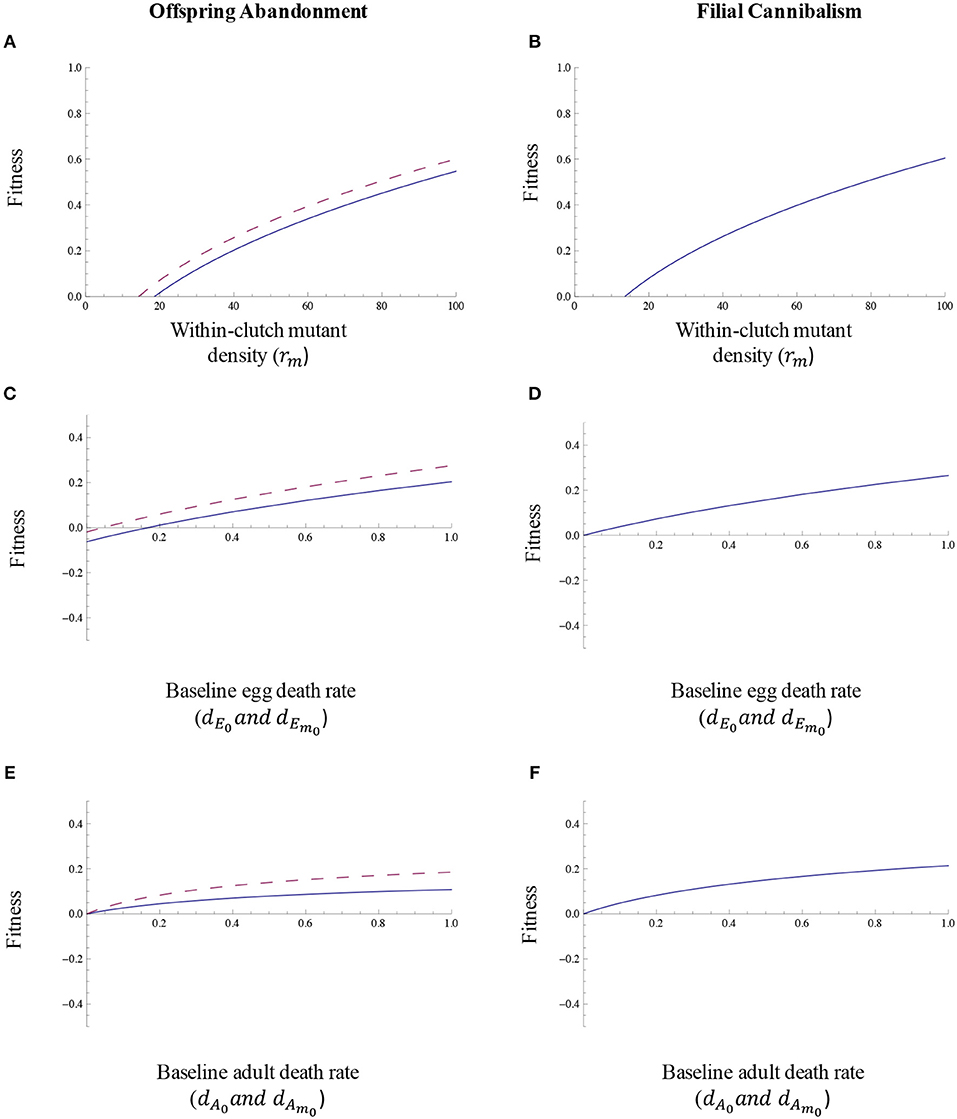
Figure 7. The relationship between (A,B) within-clutch mutant egg density, (C,D) baseline egg death rate, (E,F) baseline adult death rate and the fitness associated with offspring abandonment and filial cannibalism. In these scenarios, we assume that density effects are present and that; all other parameter values are as described in the text. For offspring abandonment, the solid line illustrates the scenario in which abandonment initially decreases offspring survival (abandonment scenario 1) and the dashed line illustrates the scenario in which abandonment initially increases offspring survival (abandonment scenario 2).
Discussion
Our results illustrate that offspring abandonment and filial cannibalism can themselves function and evolve as forms of parental care when density affects within-clutch egg mortality. As noted, density has been found to affect egg survival, and in some cases abandonment or cannibalism, in a range of species (Smith et al., 1987; Bjørnstad et al., 1999; Payne et al., 2002; Gunnarsson et al., 2006; Klug et al., 2006; Vallon et al., 2016). When within-clutch density effects do not influence egg mortality, neither offspring abandonment nor filial cannibalism are expected to result in positive fitness and evolve as forms of parental care. However, offspring abandonment or filial cannibalism could evolve due to other selective pressures. For example, energetic benefits (Rohwer, 1978; Manica, 2002) (see also Figure 5), oxygen availability (Payne et al., 2002, 2004), offspring developmental rate (Klug and Lindström, 2008), and mate competition (Okuda and Yanagisawa, 1996) can independently favor the evolution of offspring abandonment or filial cannibalism. Indeed, our findings (Figure 5) suggest that filial cannibalism can evolve in the absence of density effects if the energetic benefits to parents are sufficiently high.
Offspring abandonment and filial cannibalism are most likely to be able to evolve as forms of parental care when within-clutch density effects are relatively high. Indeed, we might not expect offspring abandonment or filial cannibalism to evolve as a form of parental care when within-clutch egg density is relatively low (Figure 7) or when abandoning or eating eggs has relatively little effect on reducing within-clutch egg density effects (Figure 6). Offspring abandonment and filial cannibalism are also most likely to be favored as forms of parental care when egg death rate is relatively high, as this is when offspring need some form of care the most, when adult death rate is relatively high (Figure 7), and when eggs are not damaged or killed by the process of abandonment (Figure 4B). These findings that abandonment and cannibalism that offset within-clutch density effects will be more likely to evolve when egg and adult mortality rates are high is consistent with our previous work (e.g., Klug and Bonsall, 2010). Classic life-history theory (e.g., Stearns, 1992) suggests that parents should provide care when offspring need it and that they should invest more heavily in current reproduction when mortality is relatively high, as this is when the potential for future reproductive opportunities are relatively low.
In general, the finding that offspring abandonment and filial cannibalism can, in theory, function as forms of parental care if they reduce within-clutch density effects and increase net offspring survival begs the question of why parents would ever produce more offspring than is optimal. Indeed, if within-clutch density effects cause overall egg survival to be relatively low, why would parents ever produce so many eggs? Our previous research on filial cannibalism and abandonment suggests that, all else being equal, parents should not overproduce eggs simply to abandon or eat them (Klug and Bonsall, 2007). When possible, parents should produce eggs at optimal densities. However, it is not always possible for parents to assess, in the most accurate way, the environment that their offspring will end up in. In some cases, factors that affect how many young can survive, including food availability, oxygen availability, diseases presence, and predation, might change in an unpredictable manner in some cases, particularly when abiotic and biotic factors vary spatially or temporally in a given location. Thus, we would expect offspring abandonment and filial cannibalism to be more likely to function as forms of parental care when environmental unpredictability makes it difficult for parents to assess and lay at optimal egg densities. Likewise, in some cases females deposit their eggs in the nests or territories of males and do not remain associated with their eggs. In such cases, which is a common pattern in fish species (Baylis, 1981), it may be difficult or impossible for females to identify an optimal density to lay at given that additional females might subsequently add eggs to the nest. Given this, we might expect that abandonment and cannibalism are more likely to function as forms of parental care in fish species in which males receive eggs from multiple females.
In general, it is important to note that offspring abandonment and filial cannibalism will not always function as forms of parental care, even when within-clutch density effects exist. Most likely, these behaviors will only function as forms of parental care under a relatively limited set of conditions: when within-clutch density effects exist, when abandoning or eating some young reduces such density effects, when offspring need care, and when eggs are laid at higher than optimal densities. To formulate empirical tests of whether offspring abandonment or filial cannibalism can function as forms of parental care, it will be key to examine the link between abandonment, cannibalism, and overall (i.e., total) offspring survival at a range of egg densities. Experimental manipulations of abandonment or cannibalism (e.g., through the simulation of these behaviors by removing eggs from a clutch) at a range of egg densities would be a particularly powerful way to explore whether abandonment and cannibalism can improve overall egg survival even in the absence of more conventional forms of parental care. For example, in fishes, insects, reptiles, and amphibians in which eggs are laid in close proximity to one another, we might expect to see (1) within-clutch density effects on egg survival and (2) a benefit of reducing offspring density through egg removal. To some extent, such a pattern was observed in the sand goby (Pomatoschistus minutus), as eggs are laid at high densities in this species and a previous study suggested that egg removal improved the overall survival of remaining young (Klug et al., 2006). Future empirical work is needed to evaluate whether offspring abandonment and filial cannibalism can improve overall offspring survival in a range of species.
To the best of our knowledge, the idea that offspring abandonment and filial cannibalism can themselves be and evolve as forms of parental care—even in the absence of more traditional care behaviors such as guarding or provisioning—is novel. While offspring abandonment and filial cannibalism are typically viewed as contradictory to parental care, our results illustrate that they might not be as contradictory as is commonly thought. Even if abandonment and cannibalism function as forms of parental care under only a limited set of circumstances, the finding that abandoning or consuming one's own offspring can be a form of care has the potential to broaden our understanding of what is considered to be parental care.
Data Accessibility
No raw data were generated, and all results are presented in the figures. Mathematica code for the model is publicly available in the Open Science Framework at the following link: https://osf.io/y4a82/.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
This material is based upon work supported by the National Science Foundation under Grant No. 1552721 (to HK). The authors acknowledge the support of the University of Tennessee at Chattanooga. Research reported in this publication was partially supported by the 2017 and 2018 Center of Excellence for Applied Computational Science competitions.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Alonso-Alvarez, C., and Velando, A. (2012). “Benefits and costs of parental care,” in Evolution of Parental Care, eds K. Mathias, P. T. Smiseth, and N. J. Royle (Oxford, UK: Oxford University Press), 40–61.
Baylis, J. R. (1981). The evolution of parental care in fishes, with reference to Darwin's rule of male sexual selection. Environ. Biol. Fishes 6, 223–251. doi: 10.1007/BF00002788
Bjørnstad, O. N., Fromentin, J.-M., Stenseth, N. C., and Gjøsæter, J. (1999). A new test for density-dependent survival: the case of coastal cod populations. Ecology 80, 1278–1288. doi: 10.2307/177074
Bonsall, M. B., and Klug, H. (2011a). Effects of among-offspring relatedness on the origins and evolution of parental care and filial cannibalism. J. Evol. Biol. 24, 1335–1350. doi: 10.1111/j.1420-9101.2011.02269.x
Bonsall, M. B., and Klug, H. (2011b). The evolution of parental care in stochastic environments. J. Evol. Biol. 24, 645–655. doi: 10.1111/j.1420-9101.2010.02203.x
Buzatto, B. A., Requena, G. S., Martins, E. G., and Machado, G. (2007). Effects of maternal care on the lifetime reproductive success of females in a neotropical harvestman. J. Anim. Ecol. 76, 937–945. doi: 10.1111/j.1365-2656.2007.01273.x
Clark, A. B., and Wilson, D. S. (1981). Avian breeding adaptations: hatching asynchrony, brood reduction, and nest failure. Q. Rev. Biol. 56, 253–277. doi: 10.1086/412316
Clutton-Brock, T. H. (1991). The Evolution of Parental Care. Princeton, NJ: Princeton University Press.
FitzGerald, G. J. (1992). Filial cannibalism in fishes: why do parents eat their offspring? Trends Ecol. Evol. 7, 7–10. doi: 10.1016/0169-5347(92)90190-M
Frederiksen, M., and Bregnballe, T. (2000). Evidence for density-dependent survival in adult cormorants from a combined analysis of recoveries and resightings. J. Anim. Ecol. 69, 737–752. doi: 10.1046/j.1365-2656.2000.00435.x
Gunnarsson, G., Elmberg, J., Sjöberg, K., Pöysä, H., and Nummi, P. (2006). Experimental evidence for density-dependent survival in mallard (Anas platyrhynchos) ducklings. Oecologia 149, 203–213. doi: 10.1007/s00442-006-0446-8
Klug, H., and Bonsall, M. B. (2007). When to care for, abandon, or eat your offspring: the evolution of parental care and filial cannibalism. Am. Natural. 170, 886–901. doi: 10.1086/522936
Klug, H., and Bonsall, M. B. (2010). Life history and the evolution of parental care. Evolution 64, 823–835. doi: 10.1111/j.1558-5646.2009.00854.x
Klug, H., and Bonsall, M. B. (2014). What are the benefits of parental care? the importance of parental effects on developmental rate. Ecol. Evol. 4, 2330–2351. doi: 10.1002/ece3.1083
Klug, H., Chin, A., and St Mary, C. M. (2005). The net effects of guarding on egg survivorship in the flagfish, Jordanella floridae. Anim. Behav. 69, 661–668. doi: 10.1016/j.anbehav.2004.05.019
Klug, H., and Lindström, K. (2008). Hurry-up and hatch: selective filial cannibalism of slower developing eggs. Biol. Lett. 4, 160–162. doi: 10.1098/rsbl.2007.0589
Klug, H., Lindströum, K., and Mary, C. M. S. (2006). Parents benefit from eating offspring: density-dependent egg survivorship compensates for filial cannibalism. Evolution 60, 2087–2095. doi: 10.1111/j.0014-3820.2006.tb01846.x
Knouft, J. H., Page, L. M., and Plewa, M. J. (2003). Antimicrobial egg cleaning by the fringed darter (Perciformes: Percidae: Etheostoma crossopterum): implications of a novel component of parental care in fishes. Proc. Royal Soc. London B. 270, 2405–2411. doi: 10.1098/rspb.2003.2501
Kölliker, M. (2007). Benefits and costs of earwig (Forficula auricularia) family life. Behav. Ecol. Sociobiol. 61, 1489–1497. doi: 10.1007/s00265-007-0381-7
Manica, A. (2002). Filial cannibalism in teleost fish. Biol. Rev. 77, 261–277. doi: 10.1017/S1464793101005905
Okuda, N., and Yanagisawa, Y. (1996). Filial cannibalism in a paternal mouthbrooding fish in relation to mate availability. Anim. Behav. 52, 307–314. doi: 10.1006/anbe.1996.0176
Otto, S. P., and and, Day, T. (2007). A Biologist's Guide to Mathematical Modeling in Ecology and Evolution. Princeton, NJ: Princeton University Press.
Payne, A. G., Smith, C., and Campbell, A. C. (2002). Filial cannibalism improves survival and development of beaugregory damselfish embryos. Proc. Royal Soc. B 269, 2095–2102. doi: 10.1098/rspb.2002.2144
Payne, A. G., Smith, C., and Campbell, A. C. (2004). A model of oxygen-mediated filial cannibalism in fishes. Ecol. Model. 174, 253–266. doi: 10.1016/j.ecolmodel.2003.09.026
Refsnider, J. M., and Janzen, F. J. (2010). Putting eggs in one basket: ecological and evolutionary hypotheses for variation in oviposition-site choice. Ann. Rev. Ecol. Evol. Syst. 41, 39–57. doi: 10.1146/annurev-ecolsys-102209-144712
Rohwer, S. (1978). Parent cannibalism of offspring and egg raiding as a courtship strategy. Am. Natural. 112, 429–440. doi: 10.1086/283284
Royle, N. J., Smiseth, P. T., and Kölliker, M. (2012). The Evolution of Parental Care. Oxford University Press. doi: 10.1093/acprof:oso/9780199692576.001.0001
Sean Doody, J., Steve Freedberg, B., and Scott Keogh, J. (2009). Communal egg-laying in reptiles and amphibians: evolutionary patterns and hypotheses. Q. Rev. Biol. 84, 229–252. doi: 10.1086/605078
Shepherd, J., and Cushing, D. (1980). A mechanism for density-dependent survival of larval fish as the basis of a stock-recruitment relationship. ICES J. Marine Sci. 39, 160–167. doi: 10.1093/icesjms/39.2.160
Simmons, L., and Parker, G. (1989). Nuptial feeding in insects: mating effort versus paternal investment. Ethology 81, 332–343. doi: 10.1111/j.1439-0310.1989.tb00778.x
Smith, G., Grenfell, B., and Anderson, R. (1987). The regulation of Ostertagia ostertagi populations in calves: density-dependent control of fecundity. Parasitology 95, 373–388. doi: 10.1017/S0031182000057814
Vallon, M., Anthes, N., and Heubel, K. U. (2016). Water mold infection but not paternity induces selective filial cannibalism in a goby. Ecol. Evol. 6, 7221–7229. doi: 10.1002/ece3.2403
Keywords: parental care, parental investment, offspring abandonment, filial cannibalism, life-history evolution
Citation: Davenport ME, Bonsall MB and Klug H (2019) Unconventional Care: Offspring Abandonment and Filial Cannibalism Can Function as Forms of Parental Care. Front. Ecol. Evol. 7:113. doi: 10.3389/fevo.2019.00113
Received: 30 May 2018; Accepted: 21 March 2019;
Published: 17 April 2019.
Edited by:
Sasha Raoul Xola Dall, University of Exeter, United KingdomReviewed by:
Anindita Bhadra, Indian Institute of Science Education and Research Kolkata, IndiaWendy Hood, Auburn University, United States
Copyright © 2019 Davenport, Bonsall and Klug. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hope Klug, aG9wZS1rbHVnQHV0Yy5lZHU=