 Juan A. Sanchez
Juan A. Sanchez Elena López-Gallego
Elena López-Gallego María J. Ramírez-Soria
María J. Ramírez-Soria- Department of Crop Protection, Biological Control and Ecosystem Services, Instituto Murciano de Investigación y Desarrollo Agrario y Alimentario, Murcia, Spain
Omnivorous mirids (Hemiptera: Miridae) are unusual as biocontrol agents, as they feed on both plants and pests. Therefore, extensive knowledge of their ecological behavior is required to maximize their predatory side and to minimize crop damage. Macrolophus pygmaeus is a known predator of small arthropods, used in European tomato crops for more than 20 years. This mirid is currently considered harmless to tomato, although some controversy remains in relation to the status of the species. The aim of this work was to investigate the benefits that M. pygmaeus provides as a predator and the likely damage as a plant feeder. The experiment was carried out in 6 experimental greenhouses in southern Spain. Two treatments, low and high M. pygmaeus populations, were assayed in a complete factorial randomized design with three replicates. Low and high M. pygmaeus populations were achieved by direct and pre-plant release methods, respectively. Tomato plants with a lower number of M. pygmaeus had a significantly higher number of whiteflies and Tuta absoluta galleries than those with a higher number of the mirid, and vice versa. A significantly higher proportion of aborted flowers and fruits was registered in greenhouses with higher M. pygmaeus numbers. Yield was also lower in greenhouses with higher mirid populations. The number of fruits harvested did not differ between treatments, but average fruit weight was significantly lower in the greenhouses with higher mirid numbers. The number of punctures attributed to M. pygmaeus on fruits were low in general and slightly higher in the treatment with more mirids. This work shows that M. pygmaeus provides both “services,” as an efficient biocontrol agent of key pests in tomato crops, and “disservices,” as it feeds on the reproductive organs of tomato plants, reducing yield. A deeper understanding of the factors that modulate the zoophytophagous response of this economically important species is needed.
Introduction
Omnivorous mirids (Hemiptera: Miridae) do not fit the ideal model of predators from a biological pest control perspective as they feed not only on prey but also on plants, thus providing both services and disservices. Their zoophytophagous and generalist behavior was one of the reasons why the biological pest control community was initially reluctant to include them in Integrated Pest Management (IPM) programs (Castañé et al., 2011). This point of view has greatly changed in recent decades with several species of mirids being successfully used in the regulation of pest populations, especially on vegetable crops. Several species of omnivorous mirids, such as dicyphines, are key predators in some vegetable crops (e.g., tomato) because of their ability to live on plants with glandular trichomes (Schuh and Slater, 1995; Sanchez and Cassis, 2018). Dicyphus cerastii Wagner, Dicyphus hesperus Knight, Dicyphus hyalinipennis Burmeister, Dicyphus tamanini Wagner, Macrolophus pygmaeus and Nesidiocoris tenuis Reuter (Hemiptera: Miridae) have been reported as effective biocontrol agents of small arthropod pests (Barnadas et al., 1998; Albajes and Alomar, 1999; Ceglarska, 1999; Carvalho and Mexia, 2000; Alomar et al., 2002; Sanchez et al., 2003, 2014; Calvo et al., 2012a,b). Some of these species (i.e., M. pygmaeus, N. tenuis, and D. hesperus) are routinely used in tomato crops for the control of whiteflies and the South American tomato moth Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) (Gillespie et al., 2007; Castañé et al., 2011; Calvo et al., 2012a,b; Urbaneja et al., 2012; Biondi et al., 2013, 2018). The potential of others, such as Dicyphus errans Wolf and Dicyphus maroccanus Wagner (Syn. Dicyphus bolivari Lindberg) (Hemiptera: Miridae) (Sanchez and Cassis, 2018), has been assayed recently against Tuta absoluta on tomato crops (Ingegno et al., 2013, 2017a,b; Abbas et al., 2014). Other species, such as Engytatus modestus Distant and Creontiades pallidus Rambur (Hemiptera: Miridae), are known to prey on phytophagous arthropods but have a more prominent plant-feeder character and are thus considered as pests (Urbaneja et al., 2001; Ferguson and Shipp, 2002). Nonetheless, the use of omnivorous mirids as predators is not free from controversy and while some species (i.e., N. tenuis) are openly considered as beneficial in some parts of the world (e.g., Canary Islands, southern Spain) (Carnero et al., 2000; Calvo and Urbaneja, 2004), they have been reported to cause some troubles in others (e.g., temperate Europe) (Vacante and Tropea-Garzia, 1994; Trottin-Caudal et al., 2006). Besides the likely subjective level of appreciation, it is undeniable that omnivorous mirids are unusual as predators and management strategies are needed that minimize the negative aspects of their phytophagous character while exploiting their predatory side (Albajes and Alomar, 1999; Sanchez, 2009).
Like many other heteropterans, mirids use a lacerate and flush feeding method, in which plant or prey tissues are liquified by digestive enzymes and the mechanical actions of stylets to facilitate their ingestion (Miles, 1972; Cohen, 1995; Wheeler, 2001). As phytophgous insects, they feed on the mesophyll of leaves and the ground tissues of stems, inflorescences, and fruits, which are more balanced in nutrients than phloem or xylem (Wheeler, 2001). Zoophytophagy in mirids is a continuous trait, going from strictly phytophagous species to essentially obligate carnivores (Miles, 1972; Wheeler, 2001; Cassis and Schuh, 2012). In this work, we refer to omnivorous as those species that feed obligatory or facultatively on plants but whose main feeding niche is carnivory. This is the case for some taxa, such as certain Macrolophus, Dicyphus, and Nesidiocoris species, that are of interest as biological control agents in vegetable crops and feed on both plants and prey, but for whom the contribution of vegetable nutrients to their fitness is generally very low in comparison with animal food (Sanchez et al., 2004; Ingegno et al., 2011). Besides, their perfomance varies greatly with the mirid species, host plant, the availability of alternative host plant, and even the plant part (McGregor et al., 2000; Perdikis and Lykouressis, 2000; Lucas and Alomar, 2001; Wheeler, 2001; Sanchez et al., 2004; Biondi et al., 2016). In the most favorable cases, plant feeding allows the insect to complete its development and reproduce but, generally, the nymphal stage lasts much longer and egg-laying is very limited in comparison to when feeding on prey (Naranjo and Gibson, 1996; Perdikis and Lykouressis, 2000; Sanchez et al., 2004, 2016; Urbaneja et al., 2005; Perdikis et al., 2007; Lykouressis et al., 2008; Ingegno et al., 2011; Mollá et al., 2014). In consequence, omnivorous mirids consume primarily prey when available and phytophagy, as well as plant injury, increases exponentially when animal food is scarce (Sanchez, 2008, 2009; Sanchez and Lacasa, 2008; Calvo et al., 2009; Arnó et al., 2010; Sanchez et al., 2016). Injuries produced by omnivorous mirids vary with the organ affected, and with the plant and mirid species. Mirids produce blemishes, scars, and deformations in tomato and zucchini, and minor injuries in melon fruits (Alomar and Albajes, 1996; Shipp and Wang, 2006; Castañé et al., 2011); feeding on gerbera blossoms may cause deformations and depreciation of flowers (Gillespie et al., 2007). The greatest impact of mirids derives from flower damage and fruit abortion because, rather than having an impact on the cosmetic appearance of the fruit, it materializes in the yield itself (Sanchez and Lacasa, 2008; Sanchez, 2009). This has prompted investigation into the factors that modulate the zoophytophagous response in omnivorous mirids and the development of management strategies to keep populations below critical levels (Alomar and Albajes, 1996; Sanchez and Lacasa, 2008; Sanchez, 2009; Sanchez et al., 2016).
Macrolophus pygmaeus has been marketed and included in programs for the control of pests in European tomato greenhouses for more than 20 years (Malausa and Trottin-Caudal, 1996; Castañé et al., 2011; van Lenteren, 2012). There have been problems with the identification of Macrolophus species and in many cases M. pygmaeus was identified as Macrolophus caliginosus (Martínez-Cascales et al., 2006a,b). In this work, we consider that the citations of M. caliginosus on tomato are quite likely misidentifications of M. pygmaeus (Castañé et al., 2013). This mirid was originally used for the control of whiteflies but it may potentially prey on other small arthropods - such as thrips, spider mites, leafminers, aphids, and lepidopterans, including the eggs and first instar of T. absoluta- (Fauvel et al., 1987; Alvarado et al., 1997; Barnadas et al., 1998; Riudavets and Castañé, 1998; Margaritopoulos et al., 2003; Arnó et al., 2009; Calvo et al., 2009; Urbaneja et al., 2009, 2012; Castañé et al., 2011). Macrolophus pygmaeus is currently considered as harmless to tomato crops (Castañé et al., 2011) nonetheless, some controversy still remains in relation to the status of the species. On the one hand, the damage it produces is considered as irrelevant in comparison to the benefits it provides in terms of pest control (Malausa and Trottin-Caudal, 1996; van Lenteren and Tommasini, 2003; Castañé et al., 2011). On the other hand, economic damage has been reported in tomato crops (Sampson, 1996; Sampson and Jacobson, 1999; Moerkens et al., 2016).
The fact is that no conclusive experiments have been carried out to quantify the impact of M. pygmaeus in tomato crops, and this issue has not been revised in a long time. The aim of this work was to investigate the trade-off between the benefits that M. pygmaeus provides as a pest control agent and the injuries it may cause to tomato crops in greenhouses.
Materials and Methods
Crop Setting
The assay was carried out in 6 experimental greenhouses of 5 × 10 m each at the IMIDA experimental station in Dolores de Pacheco, Murcia (Spain), from 14 February to 5 June 2018. Nothing was grown in these greenhouses in the latest 2 years and during this period they were freed of weeds, both inside and the immediate surroundings. The greenhouses were built of polycarbonate, with individual frontal and zenithal meshed openings for ventilation (10 × 14 threads/cm). The temperature was controlled independently in each greenhouse by the Mithra Clima System (Nutricontrol, S.L., Cartagena, Murcia); opening was activated automatically at 20°C and no heating or artificial lighting were provided. The temperature and relative humidity were recorded by the system every minute. There were four rows of 20 tomato plants in each greenhouse; the distance between plants in a row was 0.5 m, with 1 m between rows. Tomato seeds (cv. Optima, Seminis, Murcia) were planted on 1 December 2017 and seedlings 25–30 cm high were transplanted on 14 February 2018. This tomato cultivar is commonly cultivated in the area. The plants were grown in soil, and were watered and fertilized when needed by drip irrigation. Due to the small size of the greenhouses and their good ventilation, no pollinator insects were introduced. No pesticides nor fungicides were applied during the entire period of the assay.
Experimental Design and Sampling
The effect of M. pygmaeus on the regulation of the populations of the whitefly Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae), as well as other pests (e.g., T. absoluta), and its impact on reproductive organs (i.e., flowers and fruits) and yield was quantified. Two release methods for M. pygmaeus were assayed, direct and pre-release, in order to achieve low and high population levels of the mirid, respectively. Hereafter, these two treatments will be referred to as “low M. pygmaeus population” and “high M. pygmaeus population.” Each treatment was replicated three times in individual greenhouses, following an experimental randomized block design. Macrolophus pygmaeus was provided by Bioline Agroscience (Essex, France). In direct release, 320 adults of M. pygmaeus (half females and half males) were released per greenhouse. The adults of M. pygmaeus were approximately 7–10 days old. In order to improve the establishment of the mirid, the adults of M. pygmaeus to be introduced in each greenhouse were kept during the 24 h prior to their release in three muslin-walled, wooden-framed cages, each with 16 tomato plants (approx. 30–40 cm tall), at 25 ± 2°C, 60–70% RH, and a 16L:8D photoperiod. Twenty-four hours later, the 16 tomato plants of each cage, with the adults of M. pygmaeus, were transplanted in their respective greenhouses (four tomato plants per row). In pre-release, the same procedure as in direct release was used but the 320 adults of M. pygmaeus were left to multiply for 1 month before their introduction in the greenhouses. The mirids were fed periodically (every 3–4 days) on Ephestia kuehniella Zeller (Lepidoptera: Pyralidae); a total of 24 g were provided to each of the three cages during the whole period. The number of M. pygmaeus was counted on six plants per cage before they were introduced in the greenhouse, the number of M. pygmaeus (nymphs and adults) in the 16 tomato plants introduced in the greenhouse being estimated as 328 ± 18 (mean ± SE) individuals. In both treatments, the tomato plants with the mirids were transplanted in the greenhouses on 27 March 2018; in the 2 weeks following this, the tomato plants were supplemented with 5 g of E. kuehniella per greenhouse. Bemisia tabaci, provided by Koppert Spain, was introduced twice in all the greenhouses, on 13 March 2018 and 21 March 2018, at 400 adults per greenhouse on each date. Tuta absoluta was not introduced because it invariably colonizes tomato greenhouses in southern Spain (Cabello, 2009).
The nymphs and adults of M. pygmaeus and the T. absoluta galleries and adults were counted in situ every 10 days (approx.) on 15 whole tomato plants, selected at random, from the date of the first whitefly release until the end of the experiment. The B. tabaci nymphs and adults were counted in situ on one leaf from the top, middle, and bottom parts of the plant. The number of M. pygmaeus and other arthropods was also recorded on these leaves. To quantify the impact of M. pygmaeus on the reproductive organs of tomato plants, the number of flowers and of those aborted was counted in a truss from the apical part of each of the 15 sampled plants on each sampling date, starting 1 week after the release of M. pygmaeus. Besides, the number of fruits and of those that did not fructify was counted in the first available truss with no flowers, when inspecting the plants from top to bottom. Fruits were harvested on three occasions: 17 and 28 May, and 4 June 2018. On the three dates, all the fruits in each greenhouse were counted and weighed. Besides, on 28 May, 50 fruits per greenhouse were chosen at random, weighed, and scored individually for T. absoluta damage and M. pygmaeus punctures. Additionally, on 4 June, fruits were harvested individually from 20 randomly selected plants in each greenhouse and processed individually. They were classified as ripe (completely red), about to ripen (partially green), or green (completely green).
Statistical Analyses
Generalized linear mixed effect models (GLMMs) were used to compare the number of M. pygmaeus on plants, the number of T. absoluta galleries on plants, and the number of B. tabaci (nymphs and adults) on leaves between treatments (low and high M. pygmaeus populations). The date of sampling was introduced in the models as a random factor. GLMMs were also used to compare the proportions of aborted flowers and the proportions of aborted fruits on trusses. The data of the abundance of M. pygmaeus on plants and the proportion of aborted flowers were normally distributed; thus, the “lmer” function (“lme4” package) was used in the analyses (R-Development-Core-Team, 2017). The data for T. absoluta galleries on plants fitted a negative binomial distribution, thus, the “glmmPQL” function (“MASS” package) set to the negative binomial family was used to perform the analyses. The number of B. tabaci (nymphs and adults) per leaf and the proportion of aborted fruits fitted log-normal distributions and their analyses were performed using “glmmPQL” (“MASS” package) set to the Gaussian distribution with the link “log.” The total yield, number of fruits, and average fruit weight (yield divided by the number of fruits) were compared between treatments using the data obtained for all the fruits of each greenhouse picked on each of the three harvesting dates. Besides, the data from the processing of single tomato fruits at the second and third harvests were used to compare the number of T. absoluta wounds, the number of M. pygmaeus punctures, and the weight of single fruits between the two treatments. In the latter analyses, the state of fruit ripening was introduced in the models as a random factor. In all cases, with the exception of T. absoluta damage, the data were normally distributed and the “lmer” function was used to perform the analyses. In the case of T. absoluta damage, “glmmPQL” (“MASS” package), set to the Gaussian distribution with the link “log,” was used. All the statistical analyses were performed using R software (R-Development-Core-Team, 2017).
Results
Population Dynamics and Impact of M. pygmaeus on Pest Insects
Macrolophus pygmaeus reached significantly higher numbers in the greenhouses where populations were built up for 1 month before their introduction (pre-release) than in those where they were released after an acclimation of only 1 day with tomato plants (direct release) [χ2(1) = 26.3, P < 0.001] (Figure 1). In the two treatments the M. pygmaeus population increased progressively until the end of the experiment (62 days after release), reaching a maximum of 13.8 ± 1.9 and 23.8 ± 3.1 (mean ± SE) individuals per plant in the greenhouses with direct and pre-release, respectively.
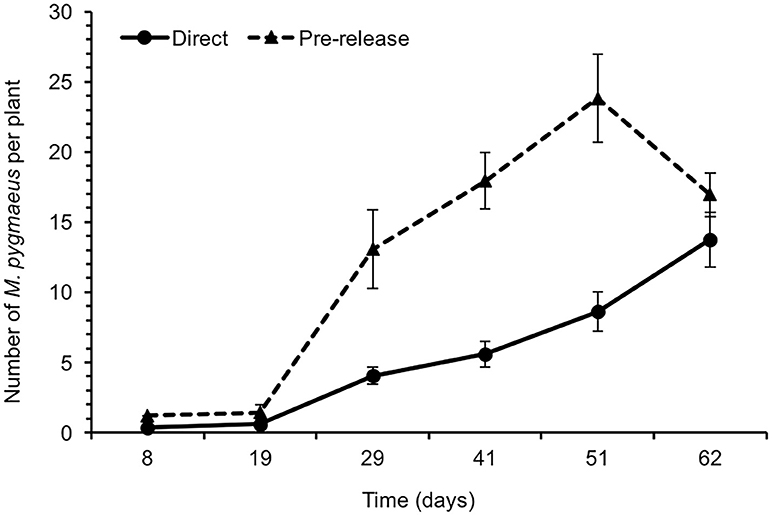
Figure 1. Average number of Macrolophus pygmaeus (adults+nymphs) per plant ± SE in tomato greenhouses with direct- and pre-release (see materials and methods). Time since the introduction of B. tabaci.
Greenhouses with a lower population of M. pygmaeus had a significantly higher number of T. absoluta galleries than those where the population of the mirid was higher [χ2(1) = 73.9, P < 0.001] (Figure 2). The number of T. absoluta galleries per plant reached a maximum of 31.0 ± 12.5 and 1.0 ± 0.6 in greenhouses with low and high M. pygmaeus populations, respectively. In the same way, the number of B. tabaci was significantly higher in greenhouses with lower rather than higher numbers of M. pygmaeus [nymphs: χ2(1) = 21.5, P < 0.001; adults: χ2(1) = 36.6, P < 0.001] (Figure 3). The whitefly numbers increased progressively in the two treatments, reaching–on day 62–a maximum of 14.8 ± 2.6 and 5.8 ± 0.6 B. tabaci nymphs per leaf in the greenhouses with lower and higher mirid density, respectively. The peak of B. tabaci adults was registered on day 41 after its introduction, being higher in greenhouses with lower (1.5 ± 0.3 adults of B. tabaci per leaf) rather than higher (0.6 ± 0.1 adults of B. tabaci per leaf) M. pygmaeus populations. No other whitefly species (T. vaporariorum), spider mites (Tetranychus spp.) or mirid (N. tenuis) were registered in samplings. Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) was observed very occassionally on leaves.
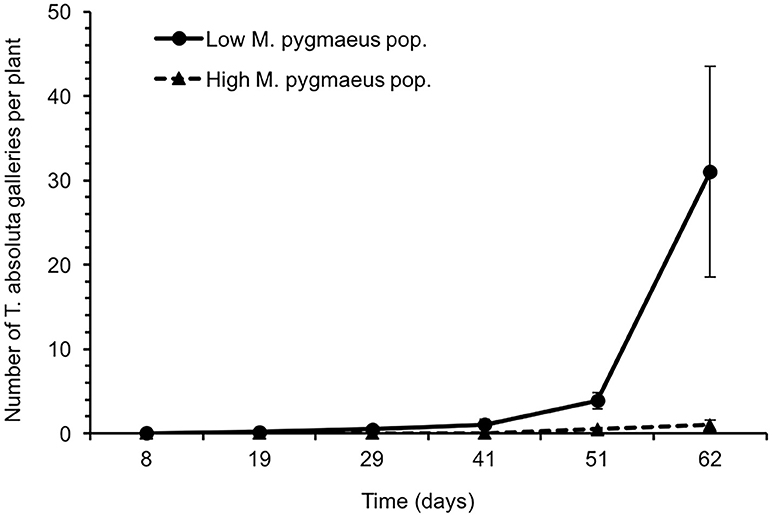
Figure 2. Average number of Tuta absoluta galleries ± SE per tomato plant in greenhouses with low and high M. pygmaeus populations. Time since the introduction of Bemisia tabaci.
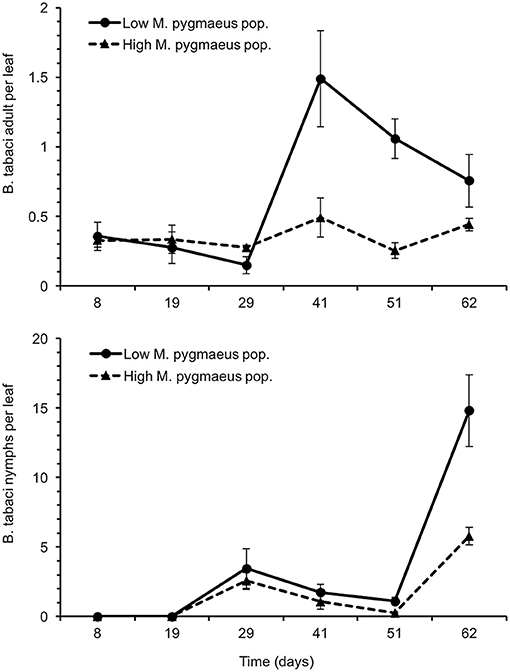
Figure 3. Average number of Bemisia tabaci nymphs (lower graph) and adults (upper graph) ± SE per leaf in tomato greenhouses with low and high M. pygmaeus populations. Time since the introduction of Bemisia tabaci.
Impact of M. pygmaeus on Reproductive Organs of Tomato Plants and Yield
A significantly higher proportion of aborted flowers was registered in greenhouses with higher M. pygmaeus numbers [χ2(1) = 15.8, P < 0.001] (Figure 4). The proportion of aborted flowers in the greenhouses with lower numbers of mirids was fairly constant and below 1.3%; in contrast, the proportion of aborted flowers reached 12.2 ± 4.1% in the greenhouses with higher mirid numbers (Figure 4). In the same way, the proportion of aborted fruits was higher in the greenhouses with higher mirid numbers [χ2(1) = 14.7, P < 0.001] (Figure 4). The percentage of aborted fruits was highest at the end of the assay, both at high (46.7 ± 9.1%) and low M. pygmaeus populations (27.9 ± 6.1%).
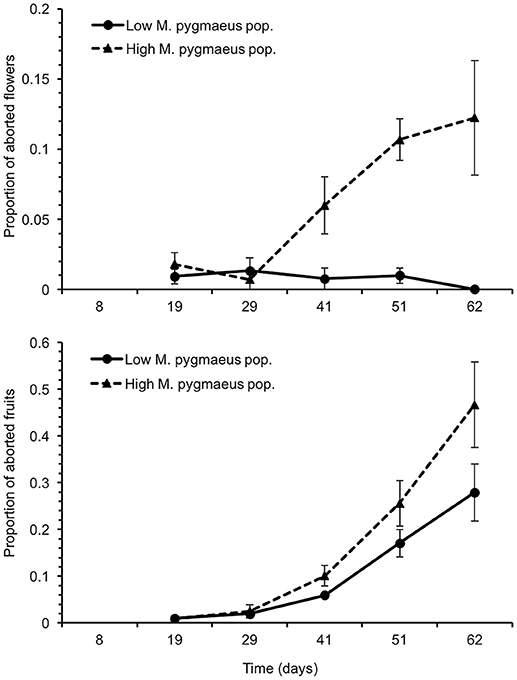
Figure 4. Proportion of aborted flowers (upper graph) and fruits (lower graph) ± SE in tomato greenhouses with low and high M. pygmaeus populations. Time since the introduction of Bemisia tabaci.
Yield was significantly lower in greenhouses with higher numbers of M. pygmaeus [χ2(1) = 4.3, P < 0.05] (Figure 5). Besides, the average fruit weight (yield/number of fruits) in the greenhouses with higher numbers of M. pygmaeus was significantly lower than in those where the mirid was less abundant [χ2(1) = 12.3, P < 0.001] (Figure 5). Similar results for fruit weight were obtained from the processing of single fruits at the second and third harvests [χ2(1) = 13.4, P < 0.001] (Supplementary Material). In contrast, the number of fruits harvested did not differ between treatments [χ2(1) = 0.114, P = 0.736]. The number of punctures attributed to M. pygmaeus was a little higher in the greenhouses with the highest densities of the mirid (mean ± SE: 1.1 ± 0.4 punctures per fruit) than in those with the lowest densities (mean ± SE: 0.8 ± 0.2 punctures per fruit) [χ2(1) = 3.1, P = 0.078] (Supplementary Material). The number of damages due to T. absoluta was higher in the greenhouses with the lowest M. pygmaeus numbers (0.053 ± 0.055 injuries per fruit) than in those with the highest numbers (0.001 ± 0.003 injuries per fruit), but not significantly so [χ2(1) = 0.153, P = 0.696] (Supplementary Material).
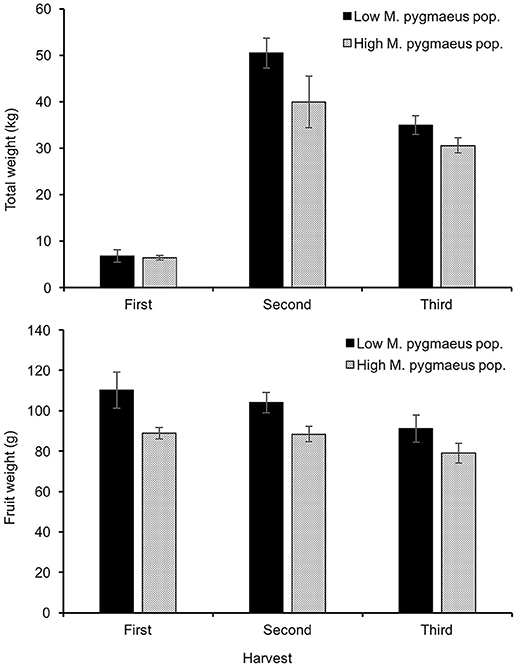
Figure 5. Total yield (upper graph) and fruit weight (lower graph) ± SE in tomato greenhouses with low and high M. pygmaeus populations.
Discussion
The two methods of introducing M. pygmaeus (pre-release and direct) in the tomato greenhouses differed in the size of the populations of the mirid that they produced. Macrolophus pygmaeus was significantly more abundant when the populations had been built up for a month before the introduction than when it was released just after a short period of acclimation (1 day) under controlled conditions. Other authors have reported differences in the establishment and population dynamics of predatory mirids depending on the release method (Lenfant et al., 2000; Calvo et al., 2012a,b; Nannini et al., 2014). In agreement with the results of this work, Nannini et al. (2014) observed that M. pygmaeus had a better establishment and reached higher population densities in tomato crops when introduced by pre-planting release, in comparison to conventional release methods.
The differences in the size of the M. pygmaeus population had opposite effects on pest control and yield. On the one hand, M. pygmaeus provided “services,” as greenhouses with higher populations of M. pygmaeus had lower incidences of two pests, T. absoluta and B. tabaci. On the other hand, it provided “disservices,” because the greenhouses with higher mirid numbers had higher flower and fruit abortion, and suffered reductions in yield and fruit size. Macrolophus pygmaeus is a reputed predator of the whiteflies B. tabaci and Trialeurodes vaporariorum Westwood (Hemiptera: Aleyrodidae) (Fauvel et al., 1987; Barnadas et al., 1998; Albajes and Alomar, 1999; Perdikis and Lykouressis, 2002; Alomar et al., 2006; Lykouressis et al., 2009; Jaeckel et al., 2011; Sylla et al., 2016). This mirid has also been reported to be one of the main predators of T. absoluta in the Mediterranean area and adjacent territories (Arnó et al., 2009; Mollá et al., 2009; Urbaneja et al., 2012; Biondi et al., 2013, 2018; Chailleux et al., 2013; Jaworski et al., 2013; Zappala et al., 2013; Sylla et al., 2016). Efficacy trials in field or semi-field conditions showed that M. pygmaeus is able to reduce the incidence and populations of T. absoluta in tomato crops (Mollá et al., 2009; Nannini et al., 2014). In the present work M. pygmaeus was able to reduce the abundance of both B. tabaci and T. absoluta; as expected, better control of T. absoluta and B. tabaci was achieved at the higher abundance of the predator. Other authors have also reported the ability of this mirid to reduce the abundance of these two pests when they were present at the same time (Bompard et al., 2013; Nannini et al., 2014). However, lower predation rates and pest control efficacy were reported for the leafminer in comparison with the whitefly (Jaworski et al., 2013; Nannini et al., 2014).
Omnivorous dicyphines are known to feed on plants, with variations in the kind of injuries they inflict on reproductive and vegetative organs, and their magnitude, depending on the mirid and plant species (Alomar and Albajes, 1996; Calvo and Urbaneja, 2004; Alomar et al., 2006; Sanchez et al., 2006; Shipp and Wang, 2006; Sanchez, 2008, 2009; Sanchez and Lacasa, 2008). Dicyphus tamaninii is known to produce small blemishes, scars, and deformation on green tomato fruits, and damage has been reported at times of high mirid abundance and low prey density (Albajes and Alomar, 1999). The feeding marks of D. hesperus were characterized as punctures surrounded by yellowish-bleached areas, and severely damaged fruit were downgraded with a 50% decrease in their market value. Shipp and Wang (2006) remarked that this mirid produced significant damage to tomato crops when its abundance was high and the prey density low, with a high proportion of severely damaged fruits when the predator:prey ratio exceeded 1:10 for D. hesperus: F. occidentalis (Shipp and Wang, 2006). Nesidiocoris tenuis is one of the species with the highest intrinsic risk of producing severe damage to tomato crops, because it produces fruit and flower abortion (Sanchez, 2008, 2009; Sanchez and Lacasa, 2008; Calvo et al., 2009), which have never been reported for any of the above mentioned dicyphines. Sanchez and Lacasa (2008) reported percentages of aborted fruits ranging from 8.79 ± 1.64 to 26.31 ± 3.53% (mean ± SE) per truss, in tomato plants exposed for 3 weeks to an average of 0.53 ± 0.26 to 35.2 ± 7.7 N. tenuis per plant, respectively. Part of this fruit abortion (10.35 ± 3.68%) was estimated to be due to causes other than N. tenuis feeding. Yield loss did not occur for the above reported range of abortion because fruit loss was compensated by an increase in the weight of individual fruit (Sanchez and Lacasa, 2008). Macrolophus pygmaeus has always been considered a safe species (Albajes and Alomar, 1999; Lucas and Alomar, 2002; Castañé et al., 2011). The injuries reported for this mirid mainly involve feeding marks on vegetative organs and tomato fruits, but they have not been generally reported in commercial settings (Malausa and Trottin-Caudal, 1996; Castañé et al., 2011). Albajes and Alomar (1999) did not observe any negative effect due to this mirid (identified as M. caliginosus) on tomato after several years of its use in conservation IPM programs; currently, it is still considered a non-problematic biocontrol agent in tomato crops (Castañé et al., 2011). Nonetheless, there is controversy in relation to the role of M. pygmaeus in tomato crops. Malausa and Trottin-Caudal (1996) found that this mirid (identified as M. caliginosus) produced damage at very high densities, even though it was considered harmless to tomato crops. Moerkens et al. (2016) argued that M. pygmaeus can cause economic feeding damage on tomato fruits at high population densities, specially when plants are infected with Pepino mosaic virus (PepMV). Sampson (1996) reported an increase in the number of fruits knuckled off in cherry tomato crops with high populations of the mirid (identified as M. caliginosus) as the prey ran short. Flower and fruit drop was reported as the most serious kind of damage to tomato crops produced by this mirid in the UK, with economic losses for both cherry and round tomato varieties (Sampson and Jacobson, 1999). The findings of the present work are in agreement with the observations of the latter authors. In the greenhouses with high M. pygmaeus populations a maximum of 12.2 ± 0.04% of flower abortion was registered, while flower abortion at low M. pygmaeus populations was always below 1.5%. Fruit abortion increased in the two treatments, but it was significantly higher at high than at low M. pygmaeus populations, with the maximum difference between treatments (18.8%) reached at the end of the assay. These differences in flower and fruit abortion between low and high M. pygmaeus populations indicates a negative impact of the mirid on tomato reproductive organs. The mirid abundance and prey availability may have both accounted for the outcome in flower and fruits abortion rates. The impact of omnivorous mirids on plant reproductive organs is known to be positively and inversely related to mirid and prey abundances, respectively (Sanchez, 2008, 2009; Sanchez and Lacasa, 2008). The much higher fruit abortion, relative to that of flowers, may have been due to intensification of M. pygmaeus feeding on young fruit petioles and/or to the effect of environmental factors. Cosmetic damage (e.g., punctures, spots) was very low and did not differ significantly between treatments. In agreement with these results, Lucas and Alomar (2002) did not find significant differences in the number of punctures between fruits exposed to the mirid (identified as M. caliginosus) and the controls. Macrolophus pygmaeus had a negative impact on production, with lower yields in greenhouses with higher M. pygmaeus populations. Surprisingly, the impact on yield was due not to the lower number of fruits at harvest but to the reduction in the weight of single fruits. This is the first time a reduction in fruit weight has been reported for any mirid species. In contrast, an increase in the weight of individual fruits that compensated fruit abortion was reported for N. tenuis (Sanchez and Lacasa, 2008; Sanchez, 2009). The reduction in fruit weight may be due to the effect of plant feeding; for instance, limited feeding of the mirid on fruit petioles could interfere with normal development without inducing fruit drop. The differences in the findings of the present work, in relation to what has been reported previously by other authors, may be due to the existence of M. pygmaeus subpopulations with different biological attributes. Ecological studies on the structure of M. pygmaeus populations using molecular markers showed the existence of several subpopulations in the Mediterranean area (Sanchez et al., 2012; Streito et al., 2017).
Whatever the reasons behind the differences between our findings and what has been reported previously for M. pygmaeus by other authors, the results of the present work raise enough concern to revise the status of this mirid. Macrolophus pygmaeus is a species of high economic importance that is currently used for pest control in more than 20 European countries and in many others in northern and southern Africa (van Lenteren, 2012). Therefore, the current consideration of this mirid as a “safe” biocontrol agent may have important economic repercussions, worldwide, for tomato crops unless proper management strategies are adopted. Establishment has always been considered a limiting factor for omnivorous mirids and researchers have always looked for strategies to establish high populations prior to pest infestations (Lenfant et al., 2000; Calvo et al., 2012a,b; Backer et al., 2014; Nannini et al., 2014). The results of the present work show that the risk of the early establishment of high populations of omnivorous mirids, M. pygmaeus included, should be taken into account. High populations may provide better pest control but that may not compensate for the yield loss derived from the increased plant feeding by the mirid. Molecular tools should be used to identify different subpopulations or strains (Sanchez et al., 2012; Streito et al., 2017), in order to select those with the most desirable biological attributes. These tools may be used also to monitor populations after their release or to detect natural immigration into crops (Sanchez et al., 2012; Streito et al., 2017). The selection of populations or strains and the implementation of appropriate management strategies will help to take advantage of the predatory side of this omnivorous mirid while minimizing the negative impact of its phytophagous side.
Author Contributions
JS designed the experiment, sampled in the field, analyzed the data, and wrote the manuscript. EL-G helped in designing the experiment, sampled in the field and digitalised the data. MP-M, LP-F, and MR-S helped with sampling in the field.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work has been funded by FEDER 14-20-03. We thank the workers at the IMIDA Torreblanca experimental station for technical assistance, and Javier Calvo and Koppert Spain for providing B. tabaci.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2018.00132/full#supplementary-material
Supplementary Data Sheet 1. Data on the processing of tomato fruits at second and third harvests. Average number of punctures attributed to Macrolophus pygmaeus, injuries of Tuta absoluta and fruit weight.
References
Abbas, S., Pérez-Hedo, M., Colazza, S., and Urbaneja, A. (2014). The predatory mirid Dicyphus maroccanus as a new potential biological control agent in tomato crops. BioControl 59, 565–574. doi: 10.1007/s10526-014-9587-6
Albajes, R., and Alomar, O. (1999). “Current and potential use of polyphagous predators,” in Integrated Pest and Disease Management in Greenhouse Crops, eds. R. Albajes, M. L. Gullino, J. C. Van Lenteren, and Y. Elad (Dordrecht: Kluwer Academic Publishers), 265–275.
Alomar, O., and Albajes, R. (1996). “Greenhouse whitefly (Homoptera: Aleyrodidae) predation and tomato fruit injury by the zoophytophagous predator Dicyphus tamaninii (Heteroptera-Miridae),” in Zoophytophagous Heteroptera: Implications for Life History and Integrated Pest Management, eds. O. Alomar, and R. N. Wiedenmann (Lanham, MD: Entomological Society of America), 155–177.
Alomar, Ò., Goula, M., and Albajes, R. (2002). Colonisation of tomato fields by predatory mirid bugs (Hemiptera: Heteroptera) in northern Spain. Agric. Ecosyst. Environ. 89, 105–115. doi: 10.1016/S0167-8809(01)00322-X
Alomar, O., Riudavets, J., and Castañé, C. (2006). Macrolophus caliginosus in the biological control of Bemisia tabaci on greenhouse melons. Biol. Control 36, 154–162. doi: 10.1016/j.biocontrol.2005.08.010
Alvarado, P., Balta, O., and Alomar, O. (1997). Efficiency of four Heteroptera as predators of Aphis gossypii and Macrosiphum euphorbiae (Hom.: Aphididae). Entomophaga 42, 215–226. doi: 10.1007/BF02769899
Arnó, J., Castañé, C., Riudavets, J., and Gabarra, R. (2010). Risk of damage to tomato crops by the generalist zoophytophagous predator Nesidiocoris tenuis (Reuter) (Hemiptera: Miridae). Bull. Entomol. Res. 100, 105–115. doi: 10.1017/S0007485309006841
Arnó, J., Sorribas, R., Prat, M., Matas, M., Pozo, C., Rodríguez, D., et al. (2009). Tuta absoluta, a new pest in IPM tomatoes in the northeast of Spain. IOBC wprs Bull. 49, 203–208.
Backer, L., de Megido, R. C., Haubruge, E., and Verheggen, F. J. (2014). Macrolophus pygmaeus (Rambur) as an efficient predator of the tomato leafminer Tuta absoluta (Meyrick) in Europe. a review. Biotechnol. Agron. Soc. Environ. 18, 536–543.
Barnadas, I., Gabarra, R., and Albajes, R. (1998). Predatory capacity of two mirid bugs preying on Bemisia tabaci. Entomol. Exp. Appl. 86, 215–219. doi: 10.1046/j.1570-7458.1998.00283.x
Biondi, A., Chailleux, A., Lambion, J., Zappalà, L., and Desneux, N. (2013). Indigenous natural enemies attacking Tuta absoluta (Lepidoptera: Gelechiidae) in Southern France. Egypt. J. Biol. Pest Control. 23, 117–121.
Biondi, A., Guedes, R. N. C., Wan, F., and Desneux, N. (2018). Ecology, worldwide spread, and management of the invasive south american tomato pinworm, Tuta absoluta: past, present, and future. Annu. Rev. Entomol. 63, 239–258. doi: 10.1146/annurev-ento-031616-034933
Biondi, A., Zappalà, L., Di Mauro, A., Tropea Garzia, G., Russo, A., Desneux, N., et al. (2016). Can alternative host plant and prey affect phytophagy and biological control by the Zoophytophagous mirid Nesidiocoris tenuis? BioControl 61, 79–90. doi: 10.1007/s10526-015-9700-5
Bompard, A., Jaworski, C. C., Bearez, P., and Desneux, N. (2013). Sharing a predator: can an invasive alien pest affect the predation on a local pest? Popul. Ecol. 55, 433–440. doi: 10.1007/s10144-013-0371-8
Cabello, T. (2009). Control biológico de la polilla del tomate, Tuta absoluta, en cultivos en invernaderos de España. Phytoma 214, 1–6.
Calvo, F. J., Bolckmans, K., and Belda, J. E. (2012a). Release rate for a pre-plant application of Nesidiocoris tenuis for Bemisia tabaci control in tomato. BioControl 57, 809–817. doi: 10.1007/s10526-012-9455-1
Calvo, F. J., Lorente, M. J., Stansly, P. A., and Belda, J. E. (2012b). Preplant release of Nesidiocoris tenuis and supplementary tactics for control of Tuta absoluta and Bemisa tabaci in greenhouse tomato. Entomol. Exp. Appl. 143, 111–119. doi: 10.1111/j.1570-7458.2012.01238.x
Calvo, F. J., and Urbaneja, A. (2004). Nesidiocoris tenuis un aliado para el control biológico de mosca blanca. Hortic. Int. 44, 20–25.
Calvo, J., Blockmans, K., Stansly, P. A., and Urbaneja, A. (2009). Predation by Nesidiocoris tenuis on Bemisia tabaci and injury to tomato. Biocontrol 54, 237–246. doi: 10.1007/s10526-008-9164-y
Carnero, A., Díaz, S., Amador, S., Hernández, M., and Hernández, E. (2000). Impact of Nesidiocoris tenuis (Heteroptera, Miridae) on whitefly populations in protected tomato crops. IOBC wprs Bull. 23:259.
Carvalho, P., and Mexia, A. (2000). First approach on the potential role of Dicyphus cerastii Wagner (Hemiptera: Miridae), as natural control agent in Portuguese greenhouses. IOBC wprs Bull. 23, 261–264.
Cassis, G., and Schuh, R. T. (2012). Systematics, biodiversity, biogeography, and host associations of the miridae (Insecta: Hemiptera: Heteroptera: Cimicomorpha). Annu. Rev. Entomol. 57, 377–404. doi: 10.1146/annurev-ento-121510-133533
Castañé, C., Agustí, N., Arnó, J., Gabarra, R., Riudavets, J., Comas, J., et al. (2013). Taxonomic identification of Macrolophus pygmaeus and Macrolophus melanotoma based on morphometry and molecular markers. Bull. Entomol. Res. 103, 204–2015. doi: 10.1017/S0007485312000545
Castañé, C., Arnó, J., Gabarra, R., and Alomar, O. (2011). Plant damage to vegetable crops by zoophytophagous mirid predators. Biol. Control 59, 22–29. doi: 10.1016/j.biocontrol.2011.03.007
Ceglarska, E. B. (1999). Dicyphus hyalinipennis Burm. (Heteroptera: Miridae): a potential biological control agent for glasshouse pests in Hungary. IOBC wprs Bull. 22, 33–36.
Chailleux, A., Bearez, P., Pizzol, J., Amiens-Desneux, E., Ramirez-Romero, R., and Desneux, N. (2013). Potential for combined use of parasitoids and generalist predators for biological control of the key invasive tomato pest Tuta absoluta. J. Pest Sci. 86, 533–541. doi: 10.1007/s10340-013-0498-6
Cohen, A. C. (1995). Extra-oral digestion in predaceous terrestrial Arthropoda. Annu. Rev. Entomol. 40, 85–103. doi: 10.1146/annurev.en.40.010195.000505
Fauvel, G., Malausa, J. C., and Kaspar, B. (1987). Laboratory studies on the main biological characteristics of Macrolophus caliginosus (Heteroptera: Miridae). Entomophaga 32, 529–543. doi: 10.1007/BF02373522
Ferguson, G., and Shipp, L. (2002). New pests in ontario greenhouse vegetables. IOBC wprs Bull. 25, 69–72.
Gillespie, D. R., McGregor, R. R., Sanchez, J. A., Vanlaerhoven, S. L., Quiring, D., Roitberg, B. D., et al. (2007). “Dicyphus hesperus (Hemiptera: Miridae) as a success story in development of endemic natural enemies as biological control agents,” in Case Studies in Biological Control: A Global Perspective, eds C. M. Goettel and G. Lazarovits (Oxfordshire, UK: CABI Publishing), 128–135.
Ingegno, B. L., Bodino, N., Leman, A., Messelink, G. J., and Tavella, L. (2017b). Predatory efficacy of Dicyphus errans on different prey. Acta Hortic. 425–430. doi: 10.17660/ActaHortic.2017.1164.55
Ingegno, B. L., Candian, V., Psomadelis, I., Bodino, N., and Tavella, L. (2017a). The potential of host plants for biological control of Tuta absoluta by the predator Dicyphus errans. Bull. Entomol. Res. 107, 340–348. doi: 10.1017/S0007485316001036
Ingegno, B. L., Ferracini, C., Gallinotti, D., Alma, A., and Tavella, L. (2013). Evaluation of the effectiveness of Dicyphus errans (Wolff) as predator of Tuta absoluta (Meyrick). Biol. Control 67, 246–252. doi: 10.1016/j.biocontrol.2013.08.002
Ingegno, B. L., Pansa, M. G., and Tavella, L. (2011). Plant preference in the zoophytophagous generalist predator Macrolophus pygmaeus (Heteroptera: Miridae). Biol. Control 58, 174–181. doi: 10.1016/j.biocontrol.2011.06.003
Jaeckel, B., Alt, S., and Balder, H. (2011). Influence of temperature and light on the feeding-rate of Macrolophus pygmaeus (Rambur, 1839) (Heteroptera: Miridae) to different whitefly species. Gesunde Pflanz. 62, 133–138. doi: 10.1007/s10343-010-0235-5
Jaworski, C. C., Bompard, A., Genies, L., Amiens-desneux, E., and Desneux, N. (2013). Preference and prey switching in a generalist predator attacking local and invasive alien pests. PLoS ONE 8:e82231. doi: 10.1371/journal.pone.0082231
Lenfant, C., Ridray, G., and Schoen, L. (2000). Biopropagation of Macrolophus caliginosus Wagner for a quicker establishment in southern tomato greenhouses. IOBC wprs Bull. 23, 247–251.
Lucas, E., and Alomar, O. (2001). Macrolophus caliginosus (Wagner) as an intraguild prey for the zoophytophagous Dicyphus tamaninii Wagner (Heteroptera: Miridae). Biol. Control 20, 147–152. doi: 10.1006/bcon.2000.0890
Lucas, E., and Alomar, O. (2002). Impact of Macrolophus caliginosus presence on damage production by Dicyphus tamaninii (Heteroptera: Miridae) on tomato fruits. J. Econ. Entomol. 95, 1123–1129. doi: 10.1603/0022-0493-95.6.1123
Lykouressis, D., Giatropoulos, A., Perdikis, D., and Favas, C. (2008). Assessing the suitability of noncultivated plants and associated insect prey as food sources for the omnivorous predator Macrolophus pygmaeus (Hemiptera: Miridae). Biol. Control 44, 142–148. doi: 10.1016/j.biocontrol.2007.11.003
Lykouressis, D. P., Perdikis, D. C., and Konstantinou, A. D. (2009). Predation rates of Macrolophus pygmaeus (Hemiptera: Miridae) on different densities of eggs and nymphal instars of the greenhouse whitefly Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Entomol. Gen. 32, 105–112. doi: 10.1127/entom.gen/32/2009/105
Malausa, J. C., and Trottin-Caudal, Y. (1996). “Advances in the strategy of use of the predaceous bug Macrolophus caliginosus (Heteroptera: Miridae) in glasshouse crops” in Zoophytophagous Heteroptera: Implications for Life History and Integrated Pest Management, eds O. Alomar and R. N. Wiedenmann (Lanham, MA: Entomological Society of America), 178–189.
Margaritopoulos, J. T., Tsitsipis, J. A., and Perdikis, D. C. (2003). Biological characteristics of the mirids Macrolophus costalis and Macrolophus pygmaeus preying on the tobacco form of Myzus persicae (Hemiptera: Aphididade). Bull. Entomol. Res. 93, 39–45. doi: 10.1079/BER2002207
Martínez-Cascales, J. I., Cenis, J. L., Cassis, G., and Sanchez, J. A. (2006a). Species identity of Macrolophus melanotoma (Costa 1853) and Macrolophus pygmaeus (Rambur 1839) (Insecta: Heteroptera: Miridae) based on morphological and molecular data and bionomic implications. Insect Syst. Evol. 37, 385–404. doi: 10.1163/187631206788831470
Martínez-Cascales, J. I., Cenis, J. L., and Sanchez, J. A. (2006b). Differentiation of Macrolophus pygmaeus (Rambur 1839) and Macrolophus melanotoma (Costa 1853) (Heteroptera: Miridae) based on molecular data. IOBC wprs Bull. 29, 223–227.
McGregor, R. R., Gillespie, D. R., Park, C. G., Quiring, D. M. J., and Foisy, M. R. J. (2000). Leaves or fruit? The potential for damage to tomato fruits by the omnivorous predator, Dicyphus hesperus. Entomol. Exp. Appl. 95, 325–328. doi: 10.1046/j.1570-7458.2000.00671.x
Miles, P. W. (1972). The saliva of hemiptera. Adv. In Insect Phys. 9, 183–255. doi: 10.1016/S0065-2806(08)60277-5
Moerkens, R., Berckmoes, E., Van Damme, V., Ortega-Parra, N., Hanssen, I., Wuytack, M., et al. (2016). High population densities of Macrolophus pygmaeus on tomato plants can cause economic fruit damage: interaction with Pepino mosaic virus? Pest Manag. Sci. 72, 1350–1358. doi: 10.1002/ps.4159
Mollá, O., Biondi, A., Alonso-Valiente, M., and Urbaneja, A. (2014). A comparative life history study of two mirid bugs preying on Tuta absoluta and Ephestia kuehniella eggs on tomato crops: implications for biological control. BioControl 59, 175–183, doi: 10.1007/s10526-013-9553-8
Mollá, O., Montón, H., Vanaclocha, P., Beitia, F., and Urbaneja, A. (2009). Predation by the mirids Nesidiocoris tenuis and Macrolophus pygmaeus on the tomato borer Tuta absoluta. IOBC wprs Bull. 49, 209–214.
Nannini, M., Atzori, F., Coinu, M., Murgia, G., Pintore, R., Pisci, R., et al. (2014). Developing improved methods for the release of Macrolophus pygmaeus (Rambur) (Heteroptera: Miridae) in Sardinian Tomato Greenhouses. Acta Hortic. 1041, 163–170. doi: 10.17660/ActaHortic.2014.1041.18
Naranjo, S. E., and Gibson, R. L. (1996). “Phytophagy in predaceous Heteroptera: effects on life history and population dynamics,” in Zoophytophagous Heteroptera: Implications for Life History and Integrated Pest Management, eds. O. Alomar, and R. N. Wiedenmann (Lanham, MA: Entomological Society of America), 57–93.
Perdikis, D., Favas, C., Lykouressis, D., and Fantinou, A. (2007). Ecological relationships between non-cultivated plants and insect predators in agroecosystems: the case of Dittrichia viscosa (Asteraceae) and Macrolophus melanotoma (Hemiptera: Miridae). Acta Oecologica 31, 299–306. doi: 10.1016/j.actao.2006.12.005
Perdikis, D., and Lykouressis, D. (2000). Effects of various items, host plants, and temperatures on the development and survival of Macrolophus pygmaeus Rambur (Hemiptera: Miridae). Biol. Control 17, 55–60. doi: 10.1006/bcon.1999.0774
Perdikis, D. C., and Lykouressis, D. P. (2002). Life table and biological characteristics of Macrolophus pygmaeus when feeding on Myzus persicae and Trialeurodes vaporariorum. Entomol. Exp. Appl. 102, 261–272. doi: 10.1046/j.1570-7458.2002.00947.x
R-Development-Core-Team (2017). A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Riudavets, J., and Castañé, C. (1998). Identification and evaluation of native predators of Frankliniella occidentalis (Thysanoptera: Thripidae) in the Mediterranean. Environ. Entomol. 27, 86–93. doi: 10.1093/ee/27.1.86
Sampson, C., and Jacobson, R. J. (1999). Macrolophus caliginosus Wagner (Heteroptera: Miridae): a predator causing damage to UK tomatoes. IOBC wprs Bull. 22, 213–216.
Sanchez, J. A. (2008). Zoophytophagy in the plantbug Nesidiocoris tenuis. Agric. For. Entomol. 10, 75–80. doi: 10.1111/j.1461-9563.2007.00357.x
Sanchez, J. A. (2009). Density thresholds for Nesidiocoris tenuis (Heteroptera: Miridae) in tomato crops. Biol. Control 51, 493–498. doi: 10.1016/j.biocontrol.2009.09.006
Sanchez, J. A., and Cassis, G. (2018). Towards solving the taxonomic impasse of the biocontrol plant bug subgenus Dicyphus (Dicyphus) (Insecta: Heteroptera: Miridae) using molecular, morphometric and morphological partitions. Zool. J. Linn. Soc. doi: 10.1093/zoolinnean/zly005. [Epub ahead of print].
Sanchez, J. A., del Amor, F. M., Flores, P., and López-Gallego, E. (2016). Nutritional variations at Nesidiocoris tenuis feeding sites and reciprocal interactions between the mirid and tomato plants. J. Appl. Entomol. 140, 161–173. doi: 10.1111/jen.12246
Sanchez, J. A., Gillespie, D. R., and McGregor, R. R. (2003). The effects of mullein (Verbascum thapsus) on the population dynamics of Dycyphus hesperus (Heteroptera: Miridae) in tomato greenhouses. Biol. Control 28, 313–319. doi: 10.1016/S1049-9644(03)00116-6
Sanchez, J. A., Gillespie, D. R., and McGregor, R. R. (2004). Plant preference in relation to life history traits in the zoophytophagous predator Dicyphus hesperus. Entomol. Exp. Appl. 112, 7–19. doi: 10.1111/j.0013-8703.2004.00174.x
Sanchez, J. A., and Lacasa, A. (2008). Impact of the zoophytophagous plant bug Nesidiocoris tenuis (Heteroptera: Miridae) on tomato yield. J. Econ. Entomol. 101, 1864–1870. doi: 10.1603/0022-0493-101.6.1864
Sanchez, J. A., La-Spina, M., and Lacasa, A. (2014). Numerical response of Nesidiocoris tenuis (Hemiptera: Miridae) preying on Tuta absoluta (Lepidoptera: Gelechiidae) in tomato crops. Eur. J. Entomol. 111, 387–395. doi: 10.14411/eje.2014.041
Sanchez, J. A., Pino-Perez, M., del Davo, M., del, Martinez-Cascales, J. I., and Lacasa, A. (2006). Zoophytophagy of the plantbug Nesidiocoris tenuis in tomato crops in southeast Spain. IOBC wprs Bull. 29, 243–248.
Sanchez, J. A., Spina, M. L., and Perera, O. P. (2012). Analysis of the population structure of Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae) in the Palaearctic region using microsatellite markers. Ecol. Evol. 2, 3145–3159. doi: 10.1002/ece3.420
Schuh, R. T., and Slater, J. A. (1995). True Bugs of the World (Hemiptera: Heteroptera). Classification and Natural History. Ithaca, NY: Comstock Publications Associates.
Shipp, J. L., and Wang, K. (2006). Evaluation of Dicyphus hersperus (Heteroptera: Miridae) for biological control of Frankliniella occidentalis (Thysanoptera: Thripidae) on greenhouse tomato. J. Econ. Entomol. 99, 414–420. doi: 10.1093/jee/99.2.414
Streito, J.-C., Clouet, C., Hamdi, F., and Gauthier, N. (2017). Population genetic structure of the biological control agent Macrolophus pygmaeus in Mediterranean agroecosystems. Insect Sci. 24, 859–876. doi: 10.1111/1744-7917.12370
Sylla, S., Brévault, T., Diarra, K., Bearez, P., and Desneux, N. (2016). Life-history traits of Macrolophus pygmaeus with different prey foods. PLoS ONE 11:e0166610. doi: 10.1371/journal.pone.0166610
Trottin-Caudal, Y., Chabriere, C., Fournier, C., Leyre, J. M., and Schoen, L. (2006). Current situation of Bemisia tabaci in protected vegetables crops in southern France. IOBC wprs Bull. 29, 53–58.
Urbaneja, A., Arán, E., Squires, P., Lara, L., and Van Der Blom, V. (2001). Aparición del chinche Creontiades pallidus Ramb. (Hemiptera: Miridae) como depredador de mosca blanca y posible causante de datos en los cultivos de pimiento en invernadero. Agrícola Vergel. 235, 396–402.
Urbaneja, A., Gonzalez-Cabrera, J., Arnó, J., and Gabarra, R. (2012). Prospects for the biological control of Tuta absoluta in tomatoes of the Mediterranean basin. Pest Manag. Sci. 68, 1215–1222. doi: 10.1002/ps.3344
Urbaneja, A., Montón, H., and Molla, O. (2009). Suitability of the tomato borer Tuta absoluta as prey for Macrolophus pygmaeus and Nesidiocoris tenuis. J. Appl. Entomol. 133, 292–296. doi: 10.1111/j.1439-0418.2008.01319.x
Urbaneja, A., Tapia, G., and Stansly, P. (2005). Influence of host plant and prey availability on developmental time and survivorship of Nesidiocoris tenius (Het.: Miridae). Biocontrol Sci. Technol. 15, 513–518. doi: 10.1080/09583150500088777
Vacante, V., and Tropea-Garzia, G. (1994). Indagini sul ruolo ecologico di Nesidiocoris tenuis (Reuter) nelle serre fredde di pomodoro del Ragusano. Inf. Fitopatol. 9, 45–48.
van Lenteren, J. C. (2012). The state of commercial augmentative biological control: plenty of natural enemies, but a frustrating lack of uptake. BioControl 57, 1–20. doi: 10.1007/s10526-011-9395-1
van Lenteren, J. C., and Tommasini, M. G. (2003). “Mass production, storage, shipment and release of natural enemies,” in Quality Control and Production of Biological Control Agents, ed. J. C. van Lenteren (Cambridge, USA: CABI Publishing).
Wheeler, A. G. (2001). Biology of the Plant Bugs (Hemiptera: Miridae). Ithaca, NY: Cornell University Press.
Keywords: omnivorous mirids, biological control, tomato pests, fruit and flower abortion, yield, economical damage
Citation: Sanchez JA, López-Gallego E, Pérez-Marcos M, Perera-Fernández LG and Ramírez-Soria MJ (2018) How Safe Is It to Rely on Macrolophus pygmaeus (Hemiptera: Miridae) as a Biocontrol Agent in Tomato Crops? Front. Ecol. Evol. 6:132. doi: 10.3389/fevo.2018.00132
Received: 15 June 2018; Accepted: 17 August 2018;
Published: 10 September 2018.
Edited by:
Alberto Pozzebon, Università degli Studi di Padova, ItalyReviewed by:
Antonio Biondi, Università degli Studi di Catania, ItalyBarbara Letizia Ingegno, Università degli Studi di Torino, Italy
Copyright © 2018 Sanchez, López-Gallego, Pérez-Marcos, Perera-Fernández and Ramírez-Soria. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan A. Sanchez, anVhbmEuc2FuY2hlejIzQGNhcm0uZXM=