 Luca Brillante
Luca Brillante Johann Martínez-Lüscher
Johann Martínez-Lüscher Runze Yu
Runze Yu Sahap Kaan Kurtural
Sahap Kaan Kurtural- 1Department of Viticulture and Enology, California State University, Fresno, Fresno, CA, United States
- 2Department of Viticulture and Enology, University of California, Davis, Davis, CA, United States
Carbon stable isotope composition of berry must at harvest (δ13C) is an integrated assessment of plant water status during grape (Vitis vinifera L.) berry ripening. Measurement of δ13C of grape juice is proposed as an alternative to traditional measurements of water status to capture the spatial variability of physiological response at the vineyard scale, i.e., zoning. We performed samplings at four different locations in California, United States, with three different cultivars of table and wine grapes (Cabernet Sauvignon, Merlot, Crimson-Seedless). Leaf physiology (photosynthesis, AN, stomatal conductance, gs) and stem water potentials (Ψstem) were routinely measured. The δ13C was measured at harvest and strong relationships were found between Ψstem (R2 = 0.71), stomatal conductance (R2 = 0.71), net carbon assimilation (R2 = 0.59) and WUEi (R2 = 0.53). The role of leaf nitrogen on the signal was assessed by evaluating relationships between leaf nitrogen and WUEi (R2 = 0.54), Ci/Ca (R2 = 0.51), δ13C (R2 = 0.44), and Ψstem (R2 = 0.37). Although nitrogen can be among the environmental factors able to affect the δ13C signal, this difference is only observable when variability in N is very large, by pooling different vineyards/varieties, but not at the within-vineyard scale. The utility of δ13C was further tested and measured on grape berries sampled on an equidistant grid in a 3.5 ha vineyard where Ψstem was also measured throughout the field season and used to delineate management zones. Physiological measurements and grape composition were correlated to soil electrical resistivity and satellite-derived vegetation index. The two management zones obtained by δ13C or Ψstem were spatially similar at 67% and allowed to separate the harvest in two pools having statistically different grape composition (soluble solids, organic acids, and anthocyanin profiles). Zoning by δ13C performed as well as zoning by Ψstem to separate grape phenolic composition, e.g., for selective harvest. Our results provided evidence that δ13C of grape must is a reliable and repeatable assessor of plant water status and gas exchange in vineyard systems that are crucial for zoning vineyards, even when irrigated, and for ground-truthing sensor maps in precision viticulture.
Introduction
The majority of the world’s viticulture areas are in arid and semi-arid regions where vineyards receive annual precipitation amounts below 700 L m–2 (Flexas et al., 2010). The growth and development of the grapevine usually correspond to the dry spring and summer months in these regions where there is little to no precipitation. The water consumption in vineyards ranges between 300 mm to 700 mm and this is greater than annual precipitation received in arid and semi-arid regions of the world (Medrano et al., 2015). Therefore, in these regions, commercial grapevine production relies on supplemental irrigation. The sustainability of grapevine production is there under threat due to growing water scarcity, rising global temperatures, and suboptimal irrigation strategies (White et al., 2006; Diffenbaugh and Scherer, 2013; Bustan et al., 2016; Bonfante et al., 2018). This problem is especially meaningful considering that grapevine is economically viable in soils with reduced fertility not well suited to other crops and that Mediterranean ecosystems adapted to grapevine are global biodiversity hotspots (Myers et al., 2000).
Agriculture is the largest user of fresh water, accounting for 99% of the global consumptive water footprint (Hoekstra and Mekonnen, 2012). This is confirmed in California, where agriculture irrigation consists of 74% of total freshwater use (Maupin et al., 2014). Under standard cultural practices in California vineyards, 140 to 220 L of irrigation water is used to produce 1 kg of wine grape (Martínez-Lüscher et al., 2017). Optimization of irrigation is not only important for environmental sustainability but also because it directly affects the yield and composition of grapes and wines (Castellarin et al., 2007; Brillante et al., 2017, 2018a). The grapevine agronomic performance under mild water deficits is well documented (Chaves et al., 2010). However, variability in the physical environment at the growing site affects grapevine water status in space and time within the same vineyard (Brillante et al., 2016a) resulting in locally inadequate irrigation within uniformly irrigated blocks (Brillante et al., 2016a, 2017) and variability problems can be exacerbated by inefficiencies in the irrigation system. This usually results in spatial differences in yield and/or grape composition at harvest (Acevedo-Opazo et al., 2008; Taylor et al., 2010; Brillante et al., 2017).
Water savings and better agronomic performances could be obtained by reducing spatial heterogeneity in plant water status (Sanchez et al., 2017), avoiding local over-irrigation, and by better tailoring irrigation strategies (Brillante et al., 2018a). Routine measurements of plant water status in the field are time-consuming and they must be constrained to few locations and time points, thus they do not easily allow a spatialized approach. Current remote sensing (Gutiérrez et al., 2018; Kustas et al., 2019) and modeling methodologies (Brillante et al., 2016b) for zoning and mapping plant water status in precision agriculture need a validation measurement that is reliable and easy to measure in multiple locations. Carbon stable isotope composition in grape musts can address this issue, as it is rapid to measure and has been proposed as a reliable continuous integrator of plant water status throughout the ripening period (Gaudillère et al., 2002). In short, the heavier stable carbon isotopes are discriminated through the difference in binary and ternary diffusivity and ribulose 1,5-diphosphate carboxylase oxygenase kinetic constants of 13CO2 and 12CO2. When stomata are closed because of water deficit this kinetic preference is reduced, as the 13CO2/12CO2 ratio increases in the sub-stomatal cavity. The resultant photoassimilates are then enriched in 13C. Water deficits are the determining factor affecting stomatal conductance and by extension the proportion of 13C assimilated (Farquhar et al., 1989). Measurements of carbon isotopic discrimination, δ13C, can be performed on different organs and growing stages for different purposes. The use of this analysis on grape musts offers operational advantages for estimating overall water status during the ripening period (Bchir et al., 2016), as it can be performed on the same substratum used for monitoring ripening, although not offering an instantaneous evaluation useful to schedule water as for the Scholander pressure chamber. Analyses of berry must δ13C can be performed once at the end of the season to characterize the spatial pattern of the water status at the field scale (Herrero-Langreo et al., 2013), or to better understand the response of the grapevine in a given vintage (Brillante et al., 2018b), or for comparing the water use efficiency of accessions in breeding programs. Although the direct relationship between δ13C and classic reporting of plant water status by leaf or stem water potential (Ψ) was shown by previous studies across the world (see Brillante et al., 2018b, for a recent meta-analysis), the relationship with gas exchange was much less investigated and when considering the measurement of δ13C on leaves contrasting results were also reported (Poni et al., 2009; Bchir et al., 2016). The relationships between grapevine predawn water potential, Ψpd and δ13C were shown to vary in the intercept across cultivars and locations, but not in the slope (Brillante et al., 2018b), offering a way to translate isotope composition values to grapevine Ψ in relative, but also rising the need for calibration in new conditions, as in California. Additional work for understanding the relationship between plant Ψ and δ13C of grape musts is needed, and also extending the study to evaluate the link between δ13C of musts and leaf gas exchange parameters that are not frequently found in the literature.
Considering the need for a measurement that can assess water status of grapevine reliably, rapidly and cost-effectively, the overarching aim of this work was to conduct a multi-area calibration of δ 13C to grapevine Ψ and leaf gas exchange parameters to better understand the δ 13C signal in viticulture, across economically important cultivars used in wine and table grape production. The specific objective of this study was to apply this knowledge to assess if intra-vineyard variability in water status and gas-exchange could be assessed by δ13C, and use δ13C to delineate management zones for selective harvest or site-specific management, or as a method to provide an effective and reliable physiological ground-truthing for sensor maps (e.g., remotely sensed vegetation indexes, soil electrical resistivity). This is crucial for vine physiology-based zoning, and therefore, to implement site-specific strategies in precision viticulture (e.g., selective harvest, design of variable-rate irrigation systems etc.).
Materials and Methods
Experimental Sites and Plant Material
The experiment was conducted during 2016 at four different experimental sites located across the state of California (Figure 1). The characteristics of these fields are reported in Table 1. In Sonoma and Galt, vineyards were planted on Cabernet-Sauvignon grafted on 110R (V. berlandieri Planch. × V. rupestris Scheele) and 1103P (V. berlandieri Planch. × V. rupestris Scheele) respectively, trained on a high-quadrilateral trellis, and spur pruned on two bilateral cordons. In Paso Robles, the vineyard was planted on Merlot Noir, grafted on 1103P, and trained on a Vertically-Shoot-Positioned (VSP) trellis and spur pruned on a unilateral cordon (see Martínez-Lüscher et al., 2019 experiment 3 for a more detailed description of this vineyard). In Delano, the vineyard was planted on Crimson-Seedless grafted on Freedom (1613-59 × Dog Ridge 5), cane pruned and on gable trellis.
Figure 1. Geographical locations of the experimental vineyards in the study.
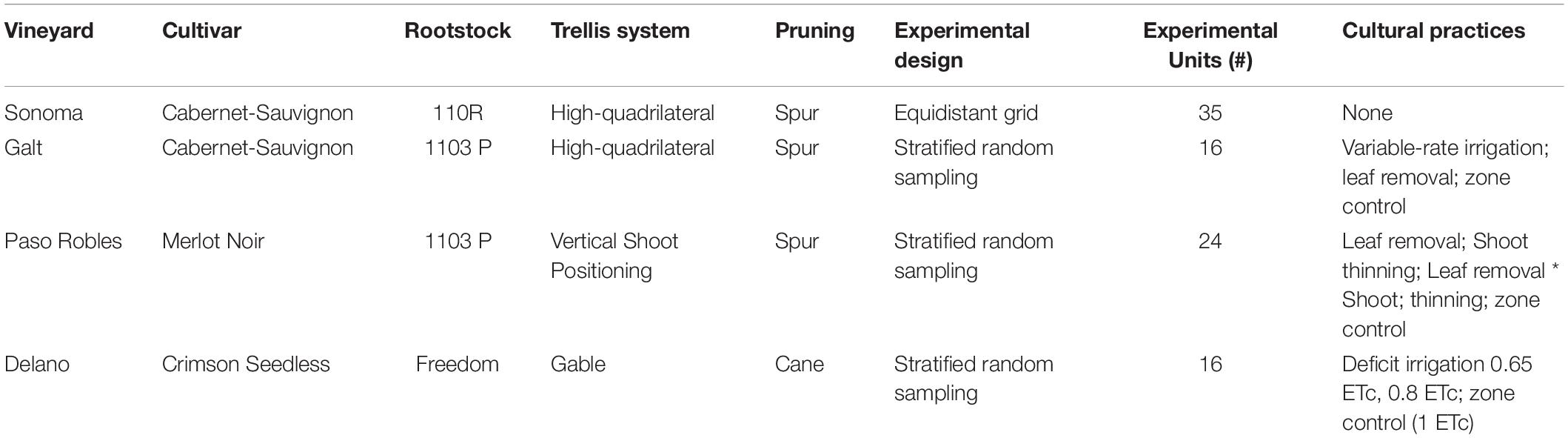
Table 1. General characteristics of the experimental design and vineyards.
Physiological measurements and berry samplings for δ13C and skin anthocyanin analysis were performed on experimental units composed of ten grapevines each and spatially distributed across each vineyard block according to a stratified random sampling based on multivariate clustering of electrical resistivity, and Normalized Difference Vegetation Index (NDVI) measured as described in the next section. In Sonoma, experimental units were instead located on a 33 m equidistant grids and composed by 5 vines each (see Brillante et al., 2017 for a more detailed description of this vineyard).
Chemical Analysis
Carbon Isotope Composition of Musts
Carbon stable isotope composition was measured in musts of mature grapes obtained from one composite sample of 100 berries, following the protocol described by Gaudillère et al. (2002). Berries were randomly sampled from multiple clusters and collected without pedicel. They were stored in ice and later crushed in the laboratory to obtain the juice. The juice was spun twice, approximately 40 ml were spun at 2000 × g and 1 ml aliquot was collected in a smaller tube and centrifuged again at 14,119 × g to further remove suspended solids. Then 5ul of the clear liquid was inserted in thin capsules, dried overnight at 60°C, and encapsulated using tweezers. Isotopic analyses were performed at the UC Davis Stable Isotope facility, using a PDZ Europa ANCA-GSL elemental analyzer interfaced to a PDZ Europa 20-20 isotope ratio mass spectrometer (Sercon Ltd., Cheshire, United Kingdom). Samples were combusted at 1080°C in a reactor packed with chromium oxide and silvered copper oxide. Following combustion, oxides were removed in a reduction reactor (reduced copper at 650°C). The helium carrier then flew through a water trap (magnesium perchlorateand phosphorous pentoxide). CO2 was retained on an adsorption trap until the N2 peak was analyzed; the adsorption trap was then heated releasing the CO2 to the IRMS. Samples were interspersed with several replicates, in addition to at least two different laboratory standards, and the standard deviation was lower than 0.2 per thousand, and conform to the long-term standard deviation of the lab. All results are expressed in delta notation, as calculated in equation (1)
where Rsample and Rstd are the absolute 13C/12C ratios for sample and standard. The values of δ13C are reported in parts per thousand respect to the Vienna Pee Dee Belemnite (VPDB) international reference.
Grape Berry Anthocyanin Extraction and Analysis
Grape berry skin anthocyanins were analyzed on 20 berries randomly sampled from each per experimental unit at maturity. Berry skins were gently peeled using a scalpel and then freeze-dried (model 7810014/7385020, Labconco, Kansas City, MO, United States). Dry skin weights were recorded after lyophilization, and then the skin tissues were powdered with a tissue lyser (MM400, Retsch, Mammelzen, Germany). Ground dry skin (50 mg) was weighed and extracted overnight at 4°C with 1 mL of methanol:water:7 M hydrochloric acid (70:29:1). Extracts were filtered with PTFE membrane filters (0.45 μm, VWR, Seattle, WA, United States), and transferred into HPLC vials before injection. The HPLC-DAD analyses of anthocyanins were performed with an Agilent 1260 (Santa Clara, CA, United States) with a LiChrospher 100, 250 mm × 4 mm with a 5 μm particle size and a 4mm guard column of the same material. HPLC gradient was the same as Ritchey and Andrew (1999).
Leaf Nitrogen Content and Plant Biomass
Vine leaf blades were collected to determine nutrient status at anthesis in each of the experimental vineyards. Fifty leaves in a position opposite a cluster were collected per experimental unit. Leaf-blades were rinsed in distilled water, dried at 65°C for 48 h and ground to pass through a 0.425 m sieve. Total nitrogen was determined by Dellavalle, Inc., Fresno, CA, United States via automated combustion analysis, method B-2.20, (Gavlak et al., 1994).
Plant biomass was estimated by collecting and weighing the wood after pruning four grapevines per experimental unit.
Supportive Analysis
Spatial Data Acquisition and Terrain Analysis
A digital elevation model was acquired using a differentially correct GPS (post-processing accuracy 2–5 cm in all directions) TRIMBLE Pro 6T DGNSS receiver (Trimble Inc., CA, United States). A terrain analysis was then performed using SAGA GIS v.2.1.2. (Conrad et al., 2015) to compute aspect, slope, and wetness index, SAGA WI (Conrad et al., 2015).
Soil Electrical Resistivity
At the beginning of the growing season, soil electrical resistivity measurements were performed using an EM38-MK2 (Geonics Limited) soil electrical conductivity meter. Measurements were performed in vertical dipole orientation at 0.75 m (Shallow ER) and 1.50 m (Deep ER). The sensor of the instrument was calibrated according to the manufacturer’s instructions to minimize the errors before the survey. The instrument was placed on a non-conductive PVC sled at an approximately 15 cm height above the ground and pulled by an all-terrain vehicle along the inter-rows at a distance of ∼2.5 m to avoid interference phenomena with the vehicle. The use of the PVC sled made it possible to keep the instrument at a constant distance from the soil surface, making data acquisition easier.
Normalized Difference Vegetation Index
At the beginning of flowering (modified E-L scale #19), canopy reflectance as NDVI was measured using a Crop Circle AS430 (Holland Scientific Inc.). The data stream was logged at 1 Hz to a GeoScout datalogger (Holland Scientific Inc.) and geolocated with a WAAS-enabled Garmin 18× GPS (Garmin Ltd.).
Solar-Noon Stem Water Potential
Plant water status was measured as stem water potentials (Ψstem). For each experimental unit and sampling instance, three (in Sonoma) to six leaves (anywhere else) from the middle section of main shoot axis were covered with a reflecting zip-top mylar bag for 2h. Around solar noon (12:30-15:30h), leaves were cut with a razor blade and immediately measured in a pressure chamber (Model 615, PMS instruments Co., OR, United States).
Leaf Gas Exchange
Leaf gas exchange was measured at solar noon (12:30-15:30h) using a portable infrared gas analyzer CIRAS-3 (PP Systems, Amesbury, MA, United States), featuring a broad-leaf chamber with 4.5 cm2 window size. In each experimental unit, three sun-exposed leaves from three grapevines and on an intermediate position of a main shoot were measured; plants and leaves randomly varied between dates. To mitigate the effect of time, the order in which experimental units were measured was randomly assigned from one measurement date to another. Assimilation rate (AN, μmol CO2 m–2 s–1) and stomatal conductance (gs, mmol H2O m–2 s–1) were obtained by measurement of inlet and outlet CO2 and H2O relative concentration. Intrinsic water use efficiency (WUEi) was calculated as the ratio between AN and gs (and then expressed in μmol CO2 mmol–1 H2O). The cuvette was oriented perpendicularly to sunlight, which was always in saturating conditions (average of internal PAR > 1900 μmol m–2 s–1). Measurements were taken at 60% relative humidity, with a CO2 concentration of 390 μmol CO2 mol–1, and using a flow to the chamber of 300 mL min–1.
Statistical Analysis
Statistical analysis was performed in R 3.5.1 (R Core Team, 2018). Generally in the text, the term significant is used to indicate p-value < 0.05.
When used, integrals of Ψstem and leaf gas exchange were calculated using the composite trapezoid rule, then divided by the time range to have values more easily comparable to individual date measurements. Correlations between parameters in the same vineyard were assessed according to a modified t-test for spatial processes (Dutilleul, 1993).
Maps of the δ13C and Ψstem were obtained by linear interpolation with x, y, z coordinates, and prediction error on unseen locations was estimated through bootstrap. Zoning was performed by k-means, and the similarity between δ13C and Ψstem zones evaluated by the Rand index.
Anthocyanin maps were performed using universal block kriging with variables of interest being linearly dependent on x, y, z coordinates, and block size 33 m ∗ 33 m. Variogram shape was assessed by multiple tests and comparisons using cross-validation. The gstat package (v. 1.1-6) was used for this purpose (Pebesma, 2004). Comparison of means across management zones was carried-out using generalized least-square ANOVA to account for spatial dependency.
The stability of the linear regression estimates (slope and R2) between δ13C and Ψstem with an increasing number of samples was assessed using a resampling routine with replacement of the whole season data set. The exercise aim was to simulate what could have happened to the δ13C ∼Ψstem relations if one or more sampling dates would have been missing from the dataset. For this purpose linear regression explaining Ψstem integrals, mean or minimum as a function of δ13C was fitted to the subset with a variable number of observations from a minimum of 3 to a maximum of total observations – 1 (6). Each resample subset was composed of unique observations and the original temporal order was conserved.
Results
Relationships Between Ψstem and Leaf Gas Exchange
A large variation was observed in the Ψstem when all cultivars (Cabernet Sauvignon, Merlot, Crimson Seedless) were pooled, in which the values ranged from −0.66 to −1.8 MPa covering a comprehensive range of plant water status for grapevine. Likewise, we also observed considerable variation in leaf gas exchange (wine grapes only). Stomatal conductance (gs) ranged from 7 to 311 mmol H2O m2 s–1, and net carbon assimilation (AN) ranged from 1.5 to 21 μmol CO2 m2 s–1. Intrinsic water use efficiency (WUEi) ranged from 0.04 to 0.29 (μmol CO2 mmol–1 H2O).
Stem water potential and leaf gas exchange parameters were linearly related across the three wine grape locations and two cultivars over which they were measured. Figure 2 presents single date averages across all experimental units. Within our range of data, the general response to an increase in Ψstem was a significant linear increase in both gs and AN. Conversely, this general response was reversed with WUEi in which as Ψstem increased, the WUEi significantly and linearly decreased. Some difference is observable between Cabernet and Merlot in the relationships of Ψsteam with gs and AN, but not with WUEi with Merlot having general lower gs and AN at equivalent Ψstem. We report these results in Supplementary Figure 1, but it is important to note that the environment was not controlled and the varieties were located in different growing regions and differentially managed, therefore limiting our ability to interpret this difference between cultivars from a physiological standpoint.
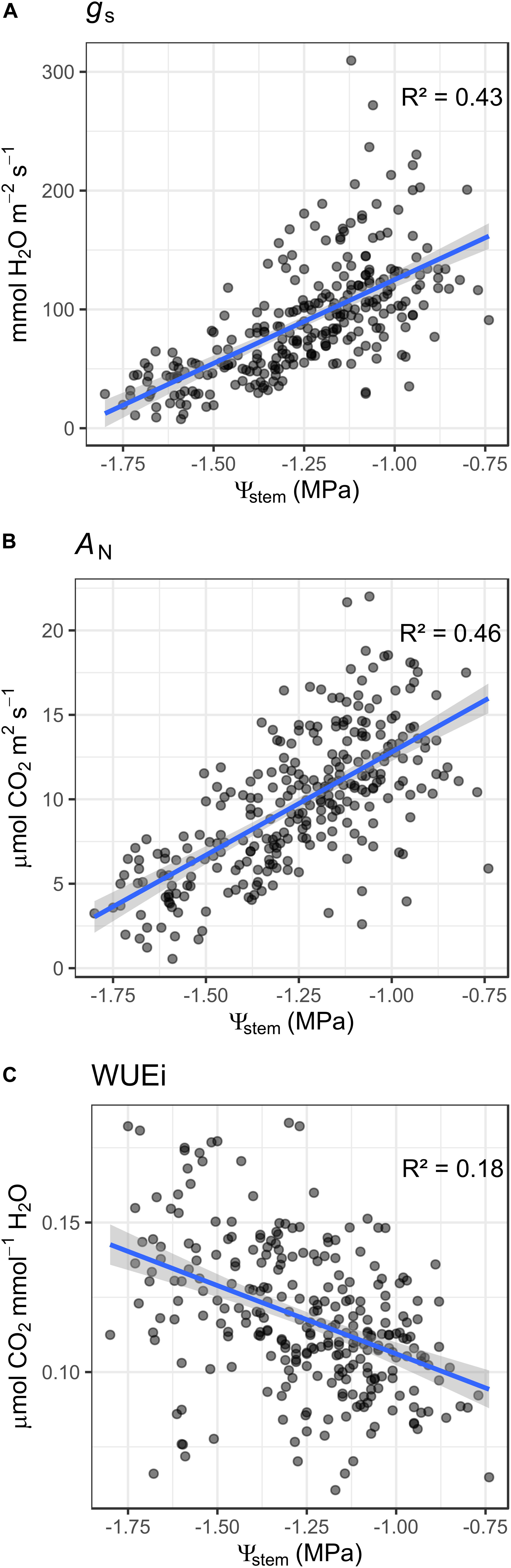
Figure 2. Relationship between stem water potential and leaf gas exchange in grapevine across three different vineyards in three different viticulture areas and two different varieties (Merlot and Cabernet-Sauvignon). Each point is an aggregate of minimum 3 measurements on different leaves and plants within a single experimental unit. (A) relationship with stomatal conductance; (gs) (B) relationship with AN (C) relationship with intrinsic water use efficiency (WUEi). Blue line is a linear regression line; in gray is the confidence interval.
Relationships Between Plant Water Status, Leaf Gas Exchange and δ13C of Grape Musts
We observed a wide range in δ13C values (−23.08 to −26.79 ‰), reflecting the variability we measured in plant water status and leaf gas exchange. The results are reported in Table 2. Compared to other δ13C values reported in the literature, values are included in the mid of the range as summarized in Brillante et al. (2018b). They show values higher (indicating less stress) than the ones reported in Gaudillère et al. (2002), for Bordeaux Cabernets and Merlot; similar to the ones reported in Guix-Hébrard et al. (2007), for Shiraz in Languedoc, FR; and lower (indicating more stress) than the ones reported in Brillante et al. (2018b) for Chardonnay in Burgundy, FR.
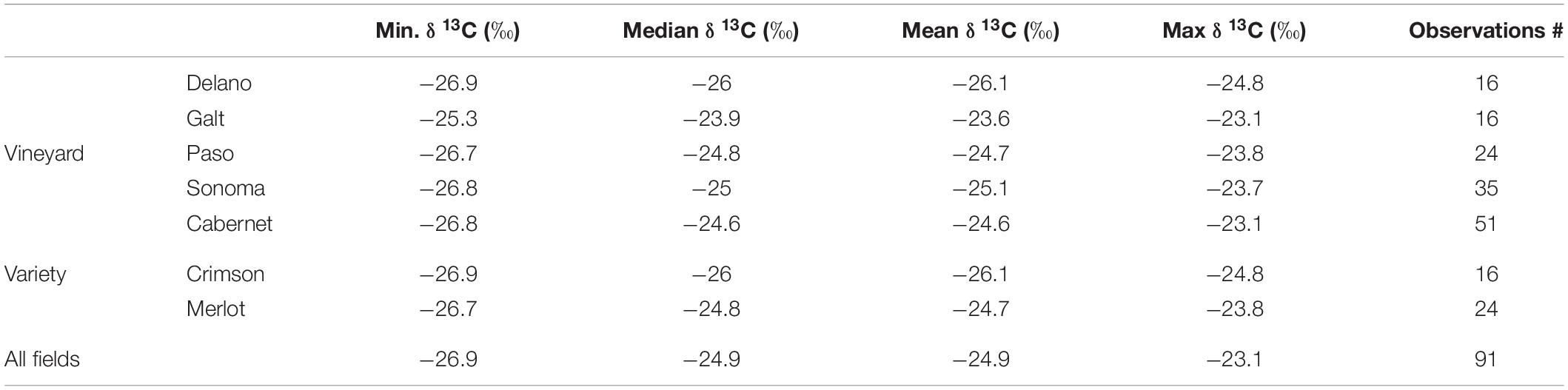
Table 2. Summary statistics of the δ13C data measured on grape juice samples
There was a strong inverse linear correlation between mean Ψstem measured across the growing season and δ13C measured on grapes at harvest, Figure 3. The relationship between leaf gas exchange (not available for Crimson Seedless) and δ13C was further confirmed by regression analysis. There was a positive relationship between season integrals of gs or AN with δ13C (Figures 4A,B). Likewise, the relationship with integrals of WUEi and δ13C was also evident, albeit in the inverse direction (Figure 4C). Supplementary Figure 2 shows differences between the two varieties in the relationships between the means of the leaf gas exchange variables and δ13C, although the intercept estimates are different the slope estimates are similar. The same limits observed in the interpretation of Supplementary Figure 1 apply here.
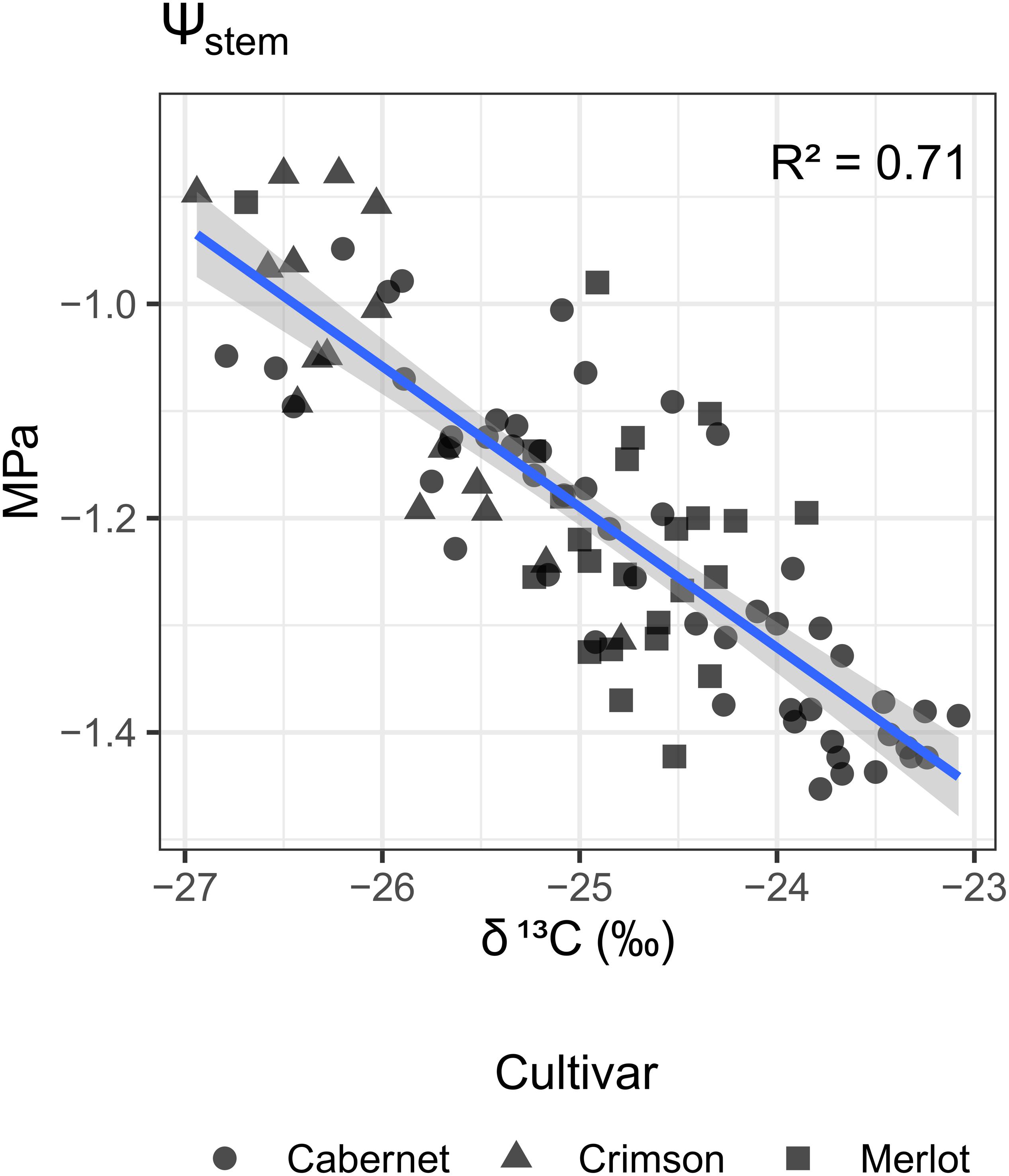
Figure 3. Relationship between δ13C of grape musts at harvest and seasonal means of Ψstem across four different vineyards in four different viticulture areas and three different varieties (Merlot, Cabernet-Sauvignon, Crimson-Seedless). Shape of point is mapped to the cultivar. Blue line is a linear regression line; in gray is the confidence interval.
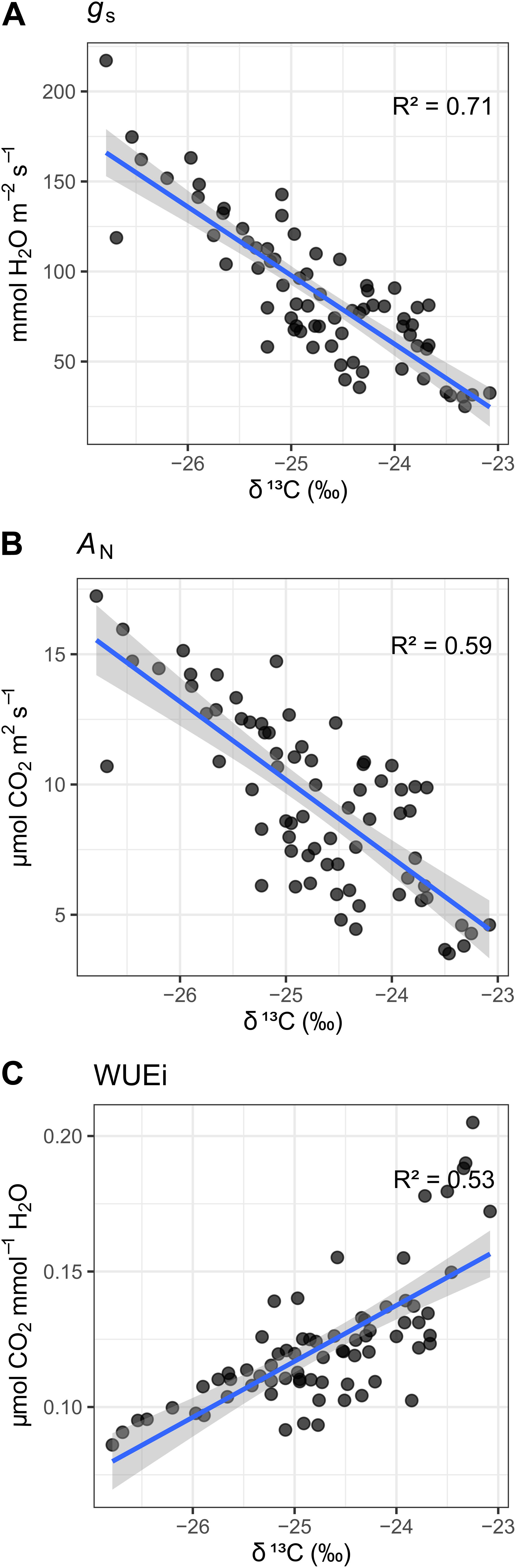
Figure 4. Relationship between δ13C of grape musts at harvest and seasonal integrals of leaf gas exchange across three different vineyards in three different viticulture areas and two different varieties (Merlot and Cabernet-Sauvignon). (A) relationship with stomatal conductance (gs), (B) 4 relationship with AN; (C) relationship with intrinsic water use efficiency (WUEi). Blue line is a linear regression line; in gray is the confidence interval. All p-values < 1e–12.
Leaf Nitrogen Concentration and Its Relationship to Gas Exchange and δ13C
Leaf nitrogen concentration was significantly and positively related to WUEi (Figure 5A). Conversely, leaf nitrogen concentration was significantly and negatively related to Ci/Ca. (Figure 5B). We also observed a significant positive relationship between leaf nitrogen concentration and δ13C (Figure 5C) when Cabernet-Sauvignon and Crimson Seedless data were pooled (nitrogen data were not available for the Merlot vineyard). The leaf nitrogen concentration was directly related to Ψstem whereas the total nitrogen concentration increased, Ψstem decreased (Figure 5C).
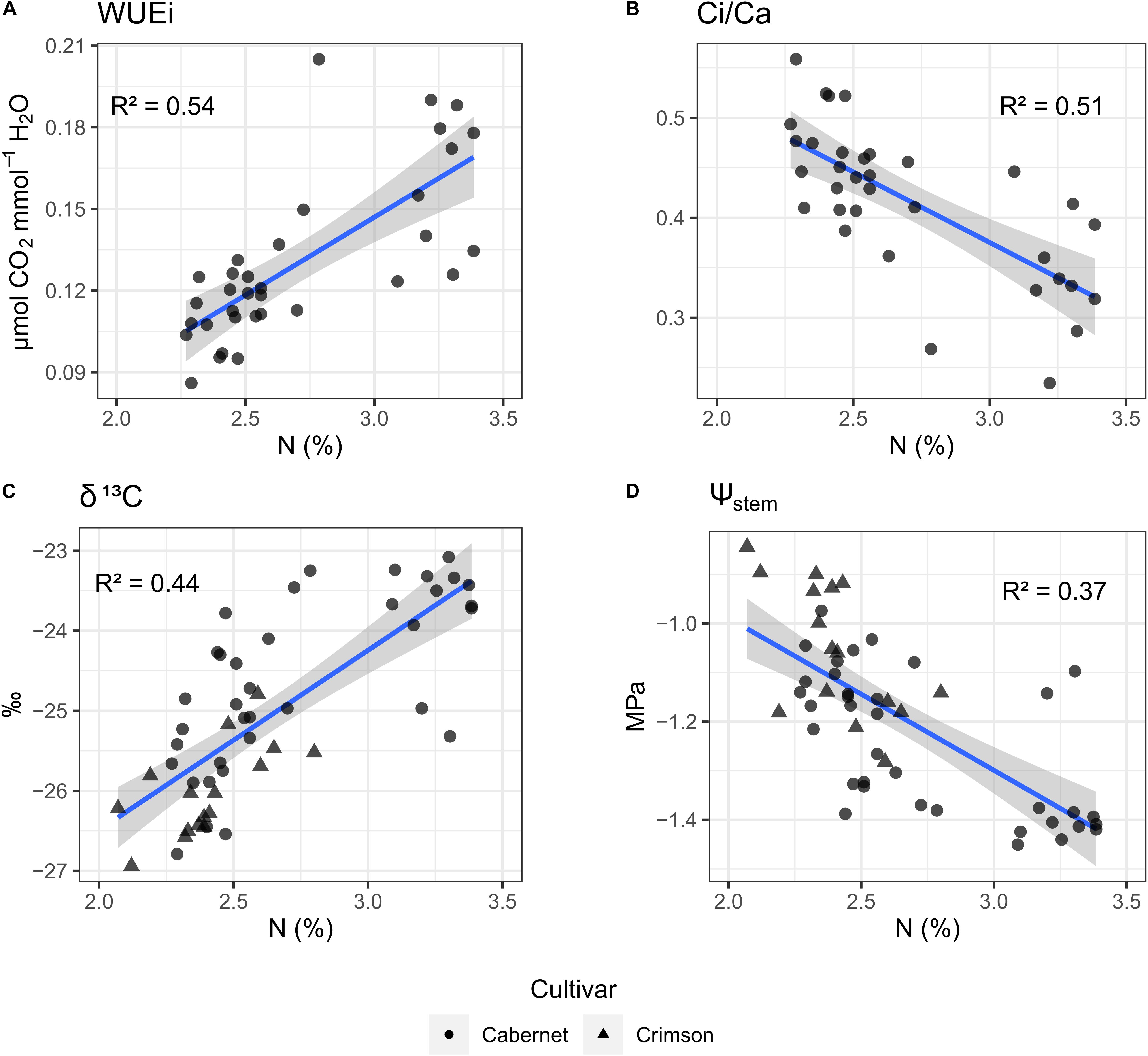
Figure 5. Relationship between Nitrogen content measured in leaf blades, and (A) seasonal integrals of WUEi, (B) seasonal integrals of Ci/Ca, (C) δ13C of musts, and (D) seasonal integrals of Ψstem. Shape of points is mapped to cultivar. Blue line is a linear regression line; in gray is the confidence interval.
Comparing δ13C and Plant Water Status to Delineate Management Zones for Selective Harvest in Precision Viticulture
Relationships Between Sensed Site Characteristics and Whole Grapevine Physiology
The physical variability of a producing vineyard was assessed by proximal sensing soil electrical resistivity, canopy reflectance, and by interpolating elevation, slope, and aspect from GPS data obtained in-situ. The shallow (0–0.75 m) and deep (0–1.50 m) soil electrical resistivity (ER), and the canopy reflectance are presented in Figure 6. In each map, experimental units where plant measurements are represented by a black circle. At this research site, soil units varied with elevation indicating a toposequence. There was a significant relationship between shallow and deep soil ER to absolute elevation measured (r = 0.73, and r = 0.70, respectively). Furthermore, soil ER was well correlated to plant water status and leaf gas exchange. It is worth noting that shallow ER (0–0.75 m) was more informative of plant conditions than deep ER (0–1.5 m). Specifically, shallow ER was significantly correlated to Ψstem, δ13C, gs, AN, and WUEi, and deep ER was significantly correlated to Ψstem only. The SAGA WI was significantly correlated to Ψstem, but not to δ13C. The NDVI was positively correlated to the topographic wetness index but not to the SAGA WI or season-long Ψstem but was positively correlated to pruned wood weight. However, wood weight was significantly and inversely correlated to δ13C, gs, and WUEi. Correlations between plant measurements and sensor data are reported in Table 3.
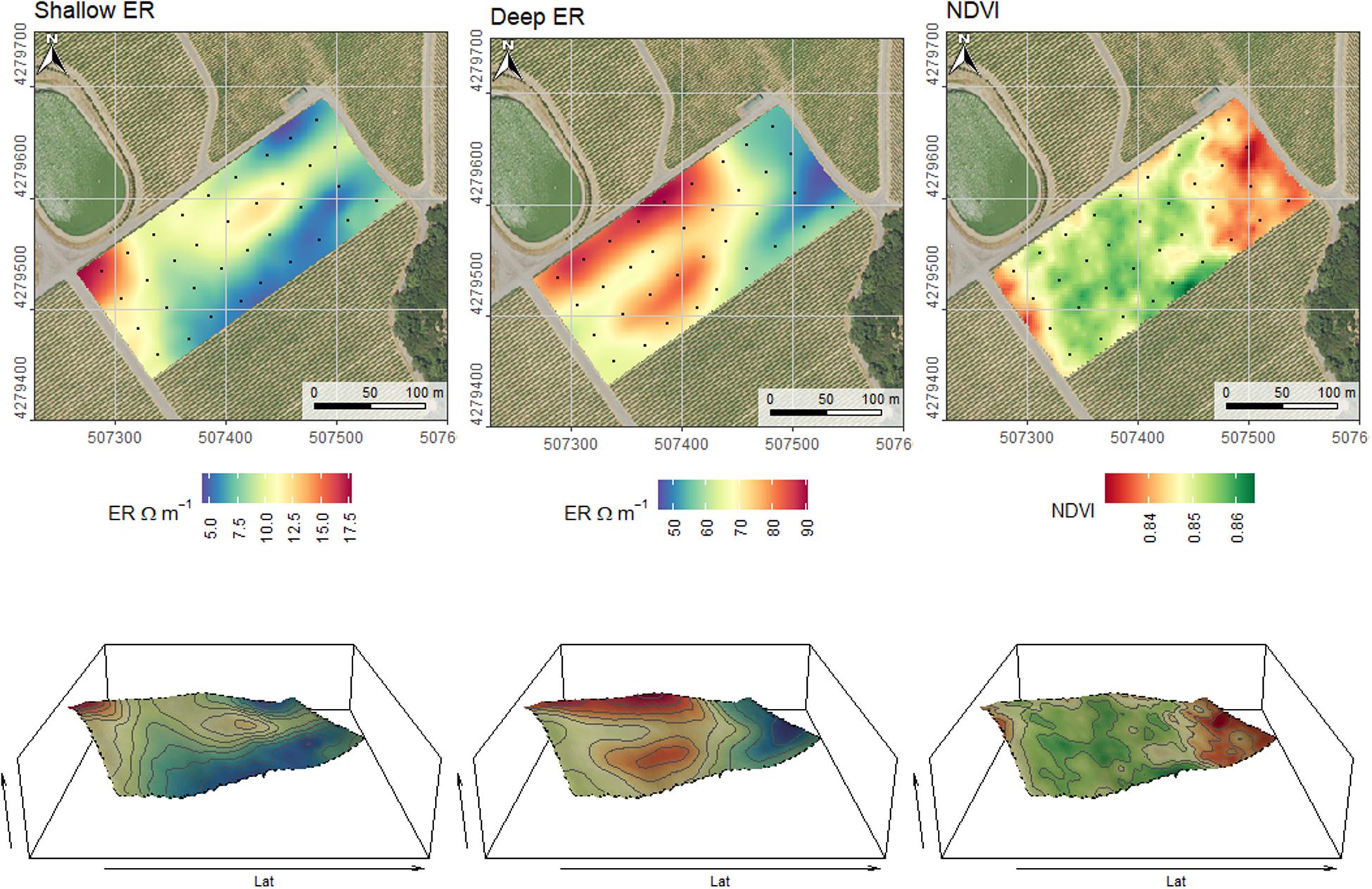
Figure 6. Maps of shallow (0–0.75 m) and deep (0–1.5 m) soil electrical resistivity and canopy reflectance as NDVI. Black dots in 2D maps shown in the upper panels indicate the locations of the experimental units. Elevation is exaggerated twice in 3D maps to enhance topographic variation within the vineyard. Coordinates are EPSG:32610 (metric).
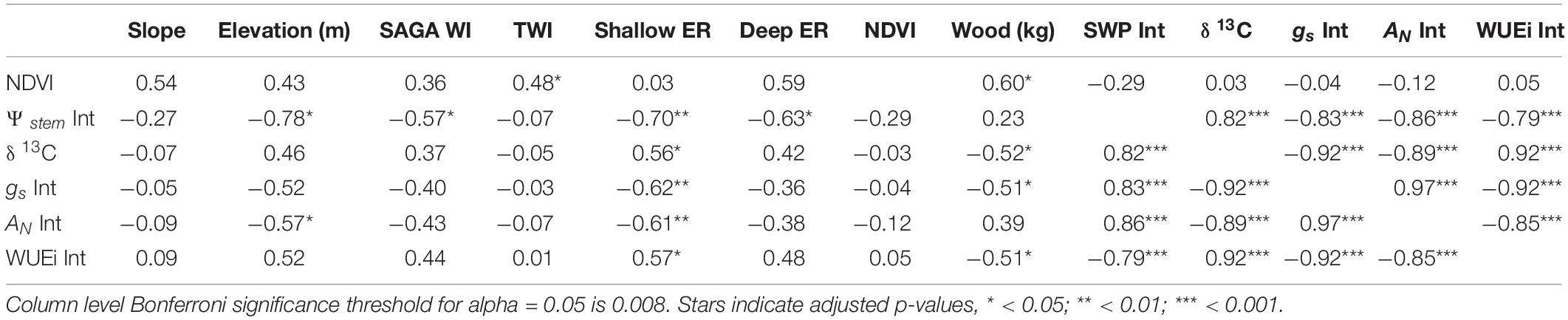
Table 3. Spatial correlations (modified t-test) between terrain characteristics, soil electrical conductivity, ground sensed NDVI data and integrals of physiological measurement of plant water potential and leaf gas exchange.
Interpolating Plant Water Status Deriving Management Zones
We interpolated plant water status by the use of Ψstem or δ13C. Figure 7A presents the map of plant water status by interpolating δ13C measured from the berries sampled in the experimental units (black dots in the figure). Figure 7B presents the map of plant water status obtained with interpolation of season-long Ψstem integrals (7 measurement dates). For δ13C the Root Mean Square Error, RMSE, in the bootstrap validation analysis was 0.63 ‰ with R2 =0.54, and for Ψstem for, the RMSE was 0.1 MPa with an R2 = 0.56. The projections of the Figures 7A,B look similar in their main trend with the North-West side of the vineyard displaying more water stress than the South-East side in either Figure. This result confirms the fundamental relation between δ13C and Ψstem shown in Figure 3, which was also established with data from this vineyard. Zoning of plant water status in two management zones for selective harvest purposes was obtained by k-means clustering applied to the underlying data of Figures 7A,B. Zones were denominated “Severe” and “Moderate” water stress, and presented in Figure 7C, δ13C derived, and 7d, Ψstem). The similarity of management zones whether calculated from Figure 7A to derive 7C or from Figure 7B to derive 7D was 0.67 as measured by the Rand index (Rand index values range between 0 and 1, for none too exact similarity).
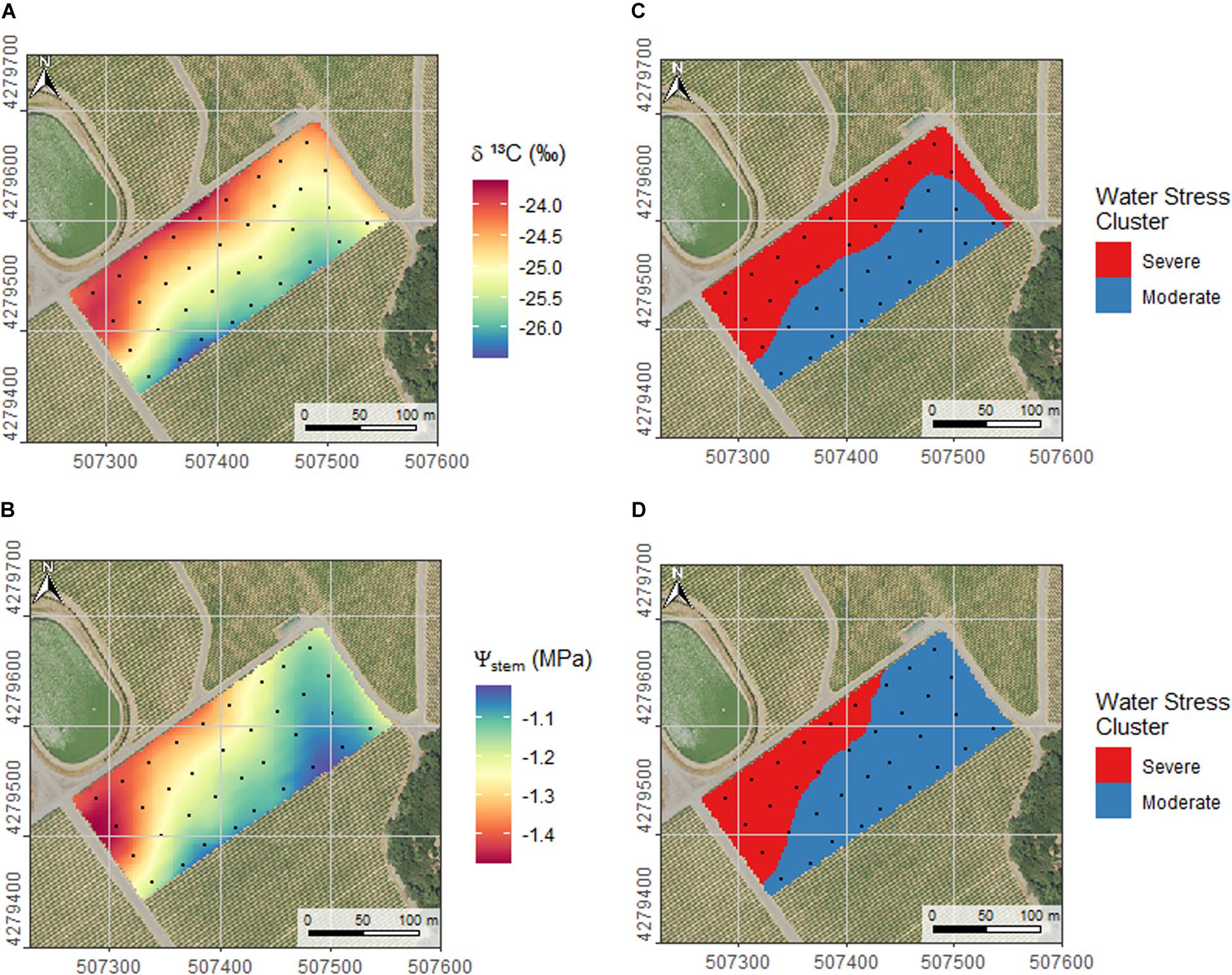
Figure 7. Interpolated maps of δ13C (A) and season integrals of Ψstem (B). Maps where interpolated using a linear interpolation with x, y and z as ancillary variables. Root mean squared error in the bootstrap validation analysis is 0.63‰ with R2 = 0.54 for interpolation of δ13C and 0.1 MPa with R2 = 0.56 for interpolation of Ψstem. K-means clustering of the field in two management zones for differential harvest according to water status throughout the season, as evaluated by δ13C (C, clustering of data in A) and Ψstem integrals (D, clustering of data in B). Similarity between the clusters in (C) and (D) as expressed by Rand index is 0.67. Coordinates are EPSG:32610 (metric).
Selective Harvest Zoning Using δ13C
Grape anthocyanin composition and profile varied greatly across the field as shown in Figure 8, and patterns closely reflected variability in water status. Assessment of variability in grape composition for selective harvest was conducted dividing the samples according to the location of experimental units within the zones in Figures 7C,D. This resulted in a total of four classes: 2 zones as separated by δ13C and 2 management zones as separated by Ψstem. The differences in anthocyanin amount and composition of grape berry as separated by these classes were analyzed with analysis of variance to compare the efficiency of δ13C or Ψstem for selective harvest. The results are presented in Figure 9. The total amount of anthocyanins, the amount of di-hydroxylated and tri-hydroxylated forms per berry basis, and the ratio between these forms was compared. The severe water stress zone had a significantly lower amount of anthocyanins on a per berry basis, a lower amount of tri-hydroxylated and of di-hydroxylated anthocyanins as well as a higher ratio between of the two hydroxylase forms compared to the moderate water stress zone. The use of δ13C or Ψstem performed, similarly, in segregating harvest zones. The preference of δ13C or Ψstem did not result in a significant difference in the anthocyanin amount or composition if one or the other plant water status assessment method was utilized.
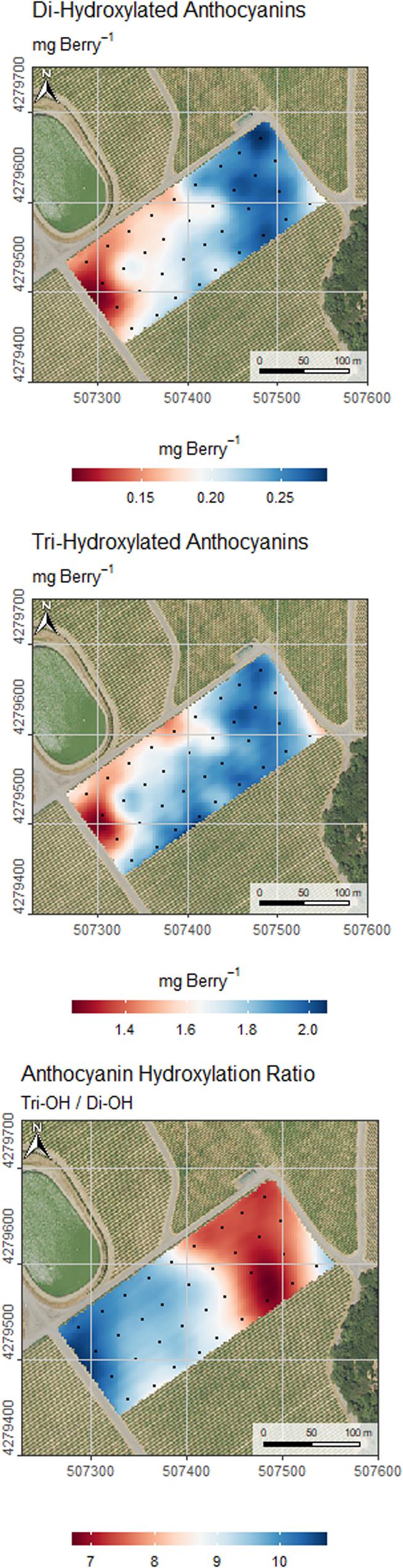
Figure 8. Kriged maps of di-hydroxylated, and tri-hydroxylated anthocyanins expressed in mg berry–1 and hydroxylation ratio (tri-hydroxylated/di-hydroxylated). Coordinates are EPSG:32610 (metric).
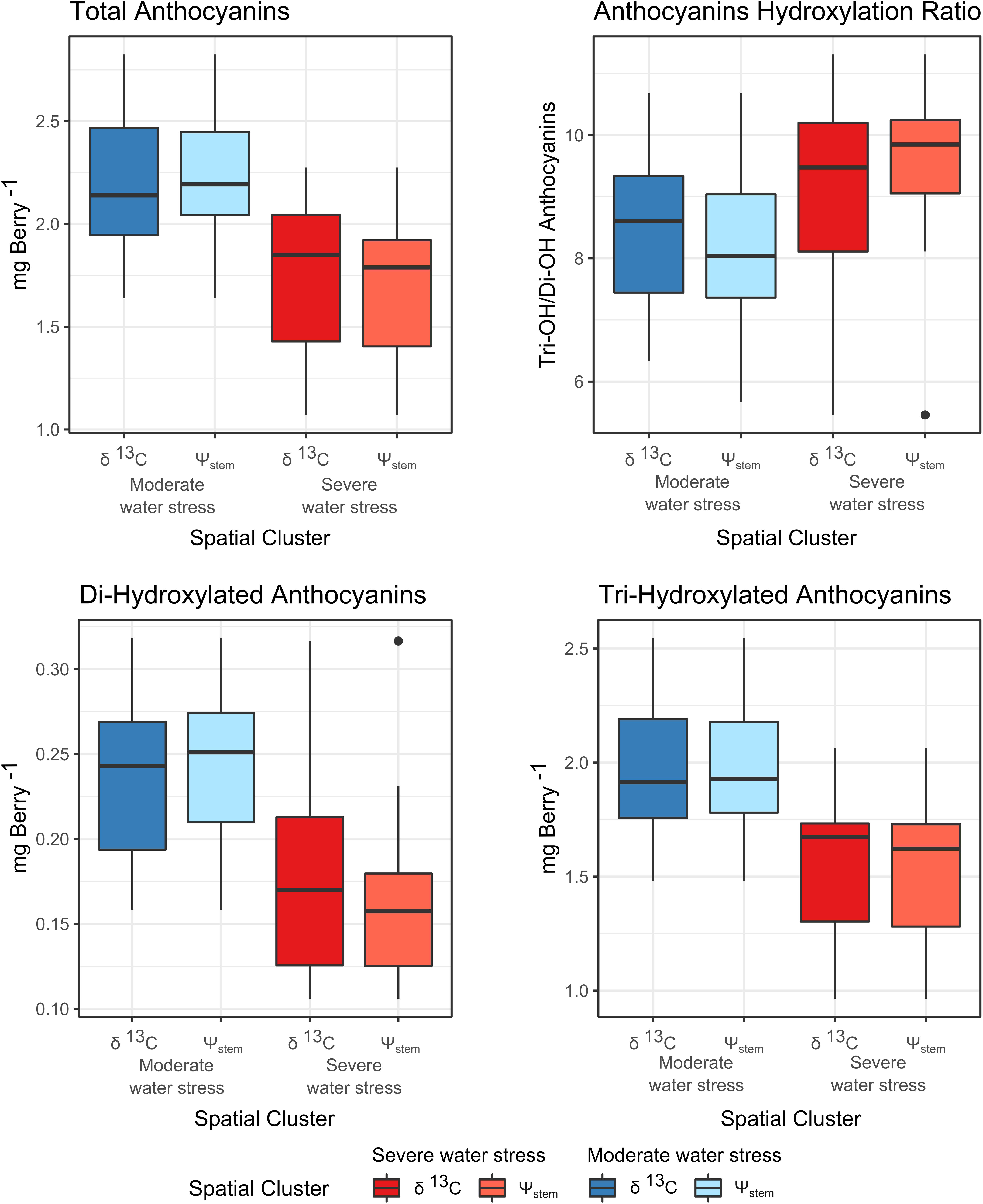
Figure 9. Separation of mean anthocyanin composition (mg berry–1) across the management zones presented in Figures 7C,D. Metabolite data from experimental units located within each zone were aggregated.
Effect of Sampling Granularity on the Correlation Between Ψstem and δ13C
The effect of sampling granularity on the regression estimates of the relations between Ψstem and δ13C of musts was assessed within the same producing Cabernet Sauvignon vineyard where the management zone clustering trial was conducted (section “Comparing δ13C and Plant Water Status to Delineate Management Zones for Selective Harvest in Precision Viticulture”). The method is described “Materials and Methods–Statistical Analysis” and results are presented in Figure 10. The purpose of this analysis was not to indicate a specific minimum number of measurements, but instead to indicate how low-quality results could be related to an insufficient sampling frequency of physiological data when compared to δ13C, which is an integrative and continuous indicator.
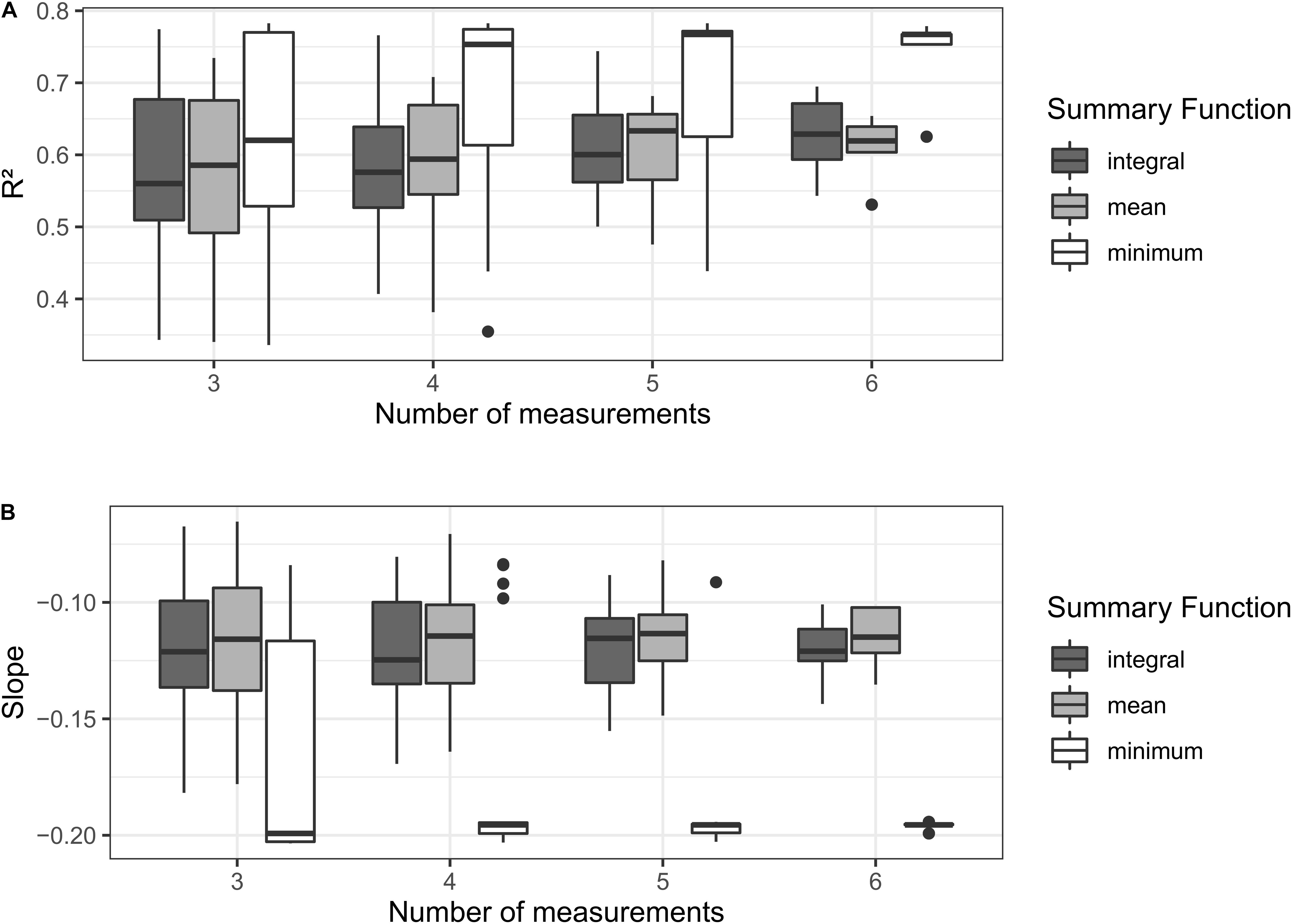
Figure 10. Evolution of the linear regression estimates between Ψstem and δ13C of musts at harvest (‰) in a simulation where measurement dates through the season for Ψstem is progressively increased. Sensitivity of different aggregation statistics for Ψstem is also tested: minimum, mean, and integral. In the linear regression Ψstem = mδ13C + b (A) shows effect on R2 and (B) shows effect on slope (m).
With the increasing number of measurement dates, the relationship between Ψstem and δ13C was stronger, low R2s were no longer estimated. The use of the minimum was able to produce the strongest relationship between variables but was the most sensitive to the number of measurements, showing the larger reduction in variability, and the larger increase in R2 with increasing frequency. This is to be expected, as by increasing the number of measurements also increases the chance of finding the same minimum value across the resample combinations. For the same reason, the minimum also had generally higher variability in R2 with respect to the use of mean or integral, except for a high number of measurements. The minimum also had lower variability in the slope with respect to the other aggregation values. A change in 1 unit of δ13C corresponds to a variation in 0.2 MPa of minimum Ψstem. The mean and the integral behaved very similarly, within the confines of our work, no clear preference for one or the other statistics could be suggested.
Discussion
Carbon Isotope Composition of Grape Musts Is a Sensitive Bio-Data Logger of Plant Water Status and Gas-Exchange During Sugar Accumulation in Berries
Determining the relationship between Ψ and leaf gas exchange parameters is necessary for physiological and agricultural interpretation of δ13C. The significant and direct relationships presented here (Figure 3) provide evidence that δ13C of grape musts was a sensitive bio-data logger of plant water status through berry development, as previously observed by other authors (Gaudillère et al., 2002; de Souza et al., 2005; Koundouras et al., 2008; Costantini et al., 2010; Brillante et al., 2018b). However, some contradictory results were also observed when δ13C was measured on leaves (Poni et al., 2009; Bchir et al., 2016). To our knowledge, our study is the first work where a relationship of δ13C of grape musts with AN and gs is presented, and between δ13C and WUEi with such a large and diverse dataset (Figure 4). Our results are corroborated by those obtained in different crops such as avocado (Persea Americana Mill., Acosta-Rangel et al., 2018), peach (Prunus persica, L., Pascual et al., 2016), and rice (Oryza sativa, L., Tao et al., 2015).
It should be kept in mind that δ13C signal is interpreted as a continuous integrator of the photosynthetic process, recording every moment of activity, while typical Ψ and leaf gas exchange determinations are discrete measurements (i.e., performed at a specific time of the day and in few time points over the growing season). Besides, when measured on sink organs such as fruits, δ13C integrates the carbon fixation processes at the scale of the whole canopy, while typical Ψ and leaf gas exchange measurements are performed on specific and selected leaves assumed representative of the whole. The differences in spatial and temporal scales of the variables in the correlations may affect the quality of the estimations. In Figure 10, we show how variability in regression estimates of the δ13C ∼ plant physiology measurements at discrete time points are affected by sampling granularity (i.e., the number of time points and summarizing function) of the discrete measurement. Not only the statistical significance and explained variability may be affected by the sampling granularity, but also slope and intercept may change. In the literature, following the seminal paper by Gaudillère et al. (2002), the use of minimum Ψ, instead than the average or the integral is frequently found to establish relationships with δ13C (Guix-Hébrard et al., 2007; Brillante et al., 2016b). The minimum Ψ ranks the worst water status conditions across the experimental units in the dataset, the correlation with δ13C appears more stable at a lower number of measurements (Figure 10) and the slope 1‰≈−0.2 MPa confirm the results of a previous meta-analysis (Brillante et al., 2018b) which was, however, based on minimum Ψpd (predawn). This correspondence δ 13C ∼Ψ will easily allow direct interpretation of relative differences in δ 13C, within and between vineyards, as absolute correspondence is not possible considering the intercept of the relationship appears to change because of environmental and genetic factors (Brillante et al., 2018b).
We would explicit that the statistical exercise shown in Figure 10 does not have the aim of identifying the minimum number of measurements to achieve reliable correlations with δ13C but to demonstrate that inconsistent results could be due to a reduced dataset for a fine integration of plant water status. We will not suggest a precise number of measurements needed for the regression analysis because this would be very dependent on the conditions of the study, as a result of the influence of weather conditions (especially vapor pressure deficit) at the time of the measurement on Ψstem, as observed in grapevine and other crops (Williams and Baeza, 2007; Suter et al., 2019). In the steady meteorological conditions of California summers, a relatively low number of measurements allows reliable correlations; in more variable weather conditions, this number could be higher. For instance, in Burgundy (Eastern France), weekly measurements of Ψstem were not related to δ13C, until a modeling approach allowed to compare averages from daily interpolations (Brillante et al., 2016b). In the same conditions, weekly measurements were enough to correlate δ13C to Ψpd (Brillante et al., 2017), as the time of the day makes it less dependent on atmospheric conditions.
Genetic and Environmental Factors and the Role of Nitrogen
In this study, we pooled together three different varieties and maintained good correlations between δ13C and other physiological measurements (Figures 3–4), but genetic variability in δ13C was reported across grapevine cultivars (Gaudillère et al., 2002; Gómez-Alonso and García-Romero, 2009; Bota et al., 2016). This variability is worth investigating as a phenotyping tool for breeding purposes, as commonly done in other crops, but it would complicate the use of δ13C in production management. The use of relative comparison as a simple turnaround has been suggested in the previous Section “Carbon Isotope Composition of Grape Musts Is a Sensitive Bio-Data Logger of Plant Water Status and Gas-Exchange During Sugar Accumulation in Berries.” The use of δ13C as a phenotyping tool would be very valuable in grapevine as well, considering the strong relationship with WUEi and gas exchange observed in Figure 4. However, several biological and environmental factors could contribute to differences in δ13C between genotypes, including stomatal behavior (Miner et al., 2017), leaf structure and anatomy (Flexas et al., 2010), and factors influencing the Ci/Ca ratio, i.e. affecting AN (Cernusak et al., 2013). In C3 species, the dependency of carbon isotope discrimination on gm has been deemed the cause of the relationships between δ13C in leaves and AN (von Caemmerer et al., 2014) which was also observed here (Figure 4). In our case, the correlation was obtained by measuring δ13C in sink organs, thus after partitioning, and not instantaneously during photosynthesis.
Comparisons of results across space and time may be complicated by genotype x environment interactions, but also by environmental conditions affecting the 13C/12C ratio in the atmosphere (e.g., latitude, altitude, etc.). Following Gaudillère et al., 2002, in grapevine literature we traditionally use δ13C, thus referring values to a chemical standard only, but the use of carbon isotope discrimination, Δ (Brook et al., 2020), thus accounting for 13C in the atmosphere, may be preferable. This approach would make data more comparable across time, considering that atmosphere is consistently enriching in 12C as a result of fossil combustion, exchange with methane, etc., and could reduce the variability in the intercept observed in Brillante et al. (2018b).
Environmental factors and interactions with horticultural practices also can play a role in variability between cultivars and complicate interpretation of results across vineyards and regions. For example, mineral nutrition affects carbon fixation rates influencing demand for CO2 in the mesophyll, Ci/Ca, and therefore, Δ (Cernusak et al., 2013). Across all mineral elements, nitrogen is the one more likely creating this effect, as shown in other woody plants such as Quercus robur L., Pinus pinaster Ait. (Guehl et al., 1995), or Ficus insipida Willd (Cernusak et al., 2007, 2013). To our knowledge, this is the first time that similar confirming results are reported in a cultivated plant such as grapevine (Figure 5). In previous work on the grapevine, where the effect of N on δ13C was also investigated (Gaudillère et al., 2002), the range of leaf N was too limited to observe any influence. In our study, a much wider range of variation was obtained by grouping data from three vineyards and two varieties; such variability is likely much harder to find in a single vineyard, in the absence of drastic variability in soil available N.
The discussion of our results in Figure 5 should consider that they were obtained under commercial production conditions and in concomitance with drought, which is likely to interact with N content, and constitute big difference respect to the previously cited works by Guehl et al. (1995) and Cernusak et al. (2007, 2013). Under the confines of our study, the correlation between Ci/Ca and N reported herein could be related to a direct increase in AN, with water stress factors also participating, as a correlation between ψ and N was observed. In principle, limited water supply is typically associated with reduced leaf expansion and lower transpiration resulting in higher leaf N content (Farquhar et al., 2002). Increased N content per leaf area has been reported as acclimation to optimize N economy under drought in willow (Salix spp.) (Weih et al., 2011), and to vary naturally in field conditions. Besides, species living in low-rainfall regions may have a greater amount of leaf N respect to species living in high rainfall environments (Wright et al., 2003). Our results corroborate previous reports in this sense, as the smallest grapevine canopies were found in water-stressed plants (as estimated by the correlation between pruned wood weights with WUEi, gs and AN in Sonoma vineyard, Table 3) having higher water use efficiency and leaf nitrogen content, while nitrogen was not correlated to the weight of dormant pruning (not shown). Our results are also in agreement with a recent meta-analysis (He and Dijkstra, 2014), indicating that availability of water, rather than N availability, could be the main driver for reduced plant growth with long-term water deficits. Future studies need to address and elucidate the inter-relationship between leaf nitrogen and carbon discrimination, across different water deficits. Although some variability in δ13C associated with N content cannot be excluded, it is unlikely that the effect of N could be so strong to prevent a direct interpretation of δ13C in terms of plant water stress, as the results of our article show. This consideration is reinforced by considering that the effect of water deficits on δ13C, and the effect of leaf N on δ13C act in the same direction, and therefore the effect of water deficits on leaf N could indirectly reinforce the signal on δ13C.
The Use of δ13C for Zoning Vineyard Variability
The variability in plant water status may influence the yield (Guilpart et al., 2014) and berry chemistry of fleshy fruits, such as grapes (Brillante et al., 2017), and originate spatial variability in agriculture performances between and within plots, as confirmed here. Although in some rain-fed premium wine regions, with a long cultivation history, this may be exploited for site-specific management purposes or to distinguish agricultural products, i.e., as one part of the terroir effect (van Leeuwen et al., 2004), in the majority of vineyards, and agricultural systems, under irrigation, this variability may be considered a problem to deal with. Having a reliable, cost and time-effective measurement of plant water status, such as the δ13C, is crucial in managing variability, across different spatial scales, from the field to the region, and for large surface modeling purposes of plant performances, such as yield (Santesteban et al., 2016). The use of δ13C to map an overall estimation of plant water status was already proposed (Herrero-Langreo et al., 2013). However, the efficiency of zoning fruit composition, compared to maps obtained through water potential measurements (Brillante et al., 2017; Gaudin et al., 2017) was never evaluated. We provide evidence that the two strategies are equivalent in their performances of delineating vineyard management zones having different anthocyanin composition (Figure 9), useful for implementing a selective harvest approach (Bramley et al., 2011). However, it is the opinion of the authors that evaluation of plant water status by direct measurement and kriging is inefficient and costly to be implemented in commercial agriculture because of the numbers of measurement points needed in space and time for a successful interpolation and estimation of vineyard conditions. The use of sensors, such as electrical resistivity or NDVI as in here, to assess variability in vineyards is straightforward and inexpensive but requires a rigorous ground-truthing to provide reliable information that could be used for informed management. For example in our case, δ13C was correlated to shallow ER, but not to deep ER or NDVI (Table 3). Measurement of δ13C in grapes is reliable and effective for this purpose, especially considering that physical limits and physiological differences between management zones are influenced by the weather of the season and as such can change across years demanding for yearly checking. Vineyard variability can be easily mapped through the use of sensors, and physiological differences between management zones obtained in this way could be assessed on a single composite berry sample for each zone. A difference in δ13C by 1‰ between the zones, corresponding to an average difference of 0.2 MPa in ψstem during the ripening period, would be enough to produce sensible effects on the grape composition, as in the case reported here.
Conclusion
Measuring carbon isotope discrimination of grape juice is fast and effective, needs to be performed only once a year for a comprehensive and reliable assessment of plant water status during the whole ripening season. The measurement is carried out on the same substratum routinely sampled to assess grape ripening and requires little additional processing. As demonstrated in this article, the relationships with grapevine gas exchange and water potentials are very robust. Noisy factors such as differences in regions, varieties, or leaf nitrogen are minimized and could be neglected when using δ13C to map variability at the within vineyard scale, but may need to be considered if the variability is expected to be large, which also depends on the space-time scale. One solution would be comparing relative and not absolute differences in δ13C values.
Carbon isotope discrimination is a continuous assessor and as such provides a more comprehensive estimate of plant physiology respect to discrete measurements obtained with other instruments or techniques allowing discrete measurements (Ψ, gas exchange). This is even more important when daily environmental variability is high. As shown here, the difference in sample granularity may affect the relationship with the discrete measurements and bring to conflicting results when the discrete measurement is aggregated from a small number of time points that do not consent a good assessment of the temporal variability.
The ability to provide reliable estimates of plant average conditions from a single measurement is an advantage of the δ13C for zoning, thus in precision viticulture, and a limit for irrigation scheduling. With the increasing availability of sensor data to monitor plant performances in agriculture fields, we need fast and reliable ground-truthing able to characterize plant physiology efficiently with a large scale and high resolution. This article demonstrated that δ13C of grape must is a reliable and repeatable assessor of grapevine water status and gas exchange in vineyard systems that are crucial for zoning vineyards, independently if irrigated, and for rapid validation of sensor maps in precision viticulture from a physiological standpoint.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
LB designed the trial, analyzed the data and wrote the first version of the manuscript. SK acquired the funding. All authors executed the trial and contributed to the final version and approved it.
Funding
The authors acknowledge the USDA-NIFA Specialty Crop Research Initiative award no. 2015-51181-24393 for funding during the execution of the study. A graduate stipend was provided to RY by the Department of Viticulture and Enology at UC Davis, Horticulture and Agronomy Graduate Group at UC Davis, and American Society for Enology and Viticulture.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor is currently organizing a Research Topic with one of the authors LB, and confirms the absence of any other collaboration.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2020.561477/full#supplementary-material
FIGURE S1 | Relationship between stem water potential and leaf gas exchange in grapevine across three different vineyards in three different viticulture areas and two different varieties (Merlot and Cabernet-Sauvignon). Each point is an aggregate of minimum 3 measurements on different leaves and plants within a single experimental unit. (a) relationship with stomatal conductance; (gs) (b) relationship with AN; (c) relationship with intrinsic water use efficiency (WUEi). Lines are linear regression line and color is mapped to cultivar; in gray is the confidence interval.
FIGURE S2 | Relationship between δ13C (‰) of grape musts at harvest and means of leaf gas exchange across three different vineyards in three different viticulture areas and two different varieties (Merlot and Cabernet-Sauvignon). (a) relationship with stomatal conductance (gs), (b) 4 relationship with AN; (c) relationship with intrinsic water use efficiency (WUEi). Blue line is a linear regression line; in gray is the confidence interval.
References
Acevedo-Opazo, C., Tisseyre, B., Guillaume, S., and Ojeda, H. (2008). The potential of high spatial resolution information to define within-vineyard zones related to vine water status. Prec. Agric. 9, 285–302. doi: 10.1007/s11119-008-9073-1
Acosta-Rangel, A., Ávila-Lovera, E., De Guzman, M. E., Torres, L., Haro, R., Lu, M., et al. (2018). Evaluation of leaf carbon isotopes and functional traits in avocado reveals water-use effi cient cultivars. Agric. Ecosyst. Environ. 263, 60–66. doi: 10.1016/j.agee.2018.04.021
Bchir, A., Escalona, J. M., Gallé, A., Hernández-montes, E., Tortosa, I., Braham, M., et al. (2016). Carbon isotope discrimination (δ13C) as an indicator of vine water status and water use efficiency (WUE): looking for the most representative sample and sampling time. Agric. Water Manag. 167, 11–20. doi: 10.1016/j.agwat.2015.12.018
Bonfante, A., Monaco, E., Langella, G., Mercogliano, P., Bucchignani, E., Manna, P., et al. (2018). A dynamic viticultural zoning to explore the resilience of terroir concept under climate change. Sci. Total Environ. 624, 294–308. doi: 10.1016/j.scitotenv.2017.12.035
Bota, J., Tomás, M., Flexas, J., Medrano, H., and Escalona, J. M. (2016). Differences among grapevine cultivars in their stomatal behavior and water use efficiency under progressive water stress. Agric. Water Manag. 164, 91–99. doi: 10.1016/j.agwat.2015.07.016
Bramley, R. G. V., Ouzman, J., and Thornton, C. (2011). Selective harvesting is a feasible and profitable strategy even when grape and wine production is geared towards large fermentation volumes. Aust. J. Grape Wine Res. 17, 298–305. doi: 10.1111/j.1755-0238.2011.00151.x
Brillante, L., Bois, B., Lévêque, J., and Mathieu, O. (2016a). Variations in soil-water use by grapevine according to plant water status and soil physical-chemical characteristics-A 3D spatio-temporal analysis. Eur. J. Agron. 77, 122–135. doi: 10.1016/j.eja.2016.04.004
Brillante, L., Martínez-Lüscher, J., and Kurtural, S. K. (2018a). Applied water and mechanical canopy management affect berry and wine phenolic and aroma composition of grapevine (Vitis vinifera L., cv. Syrah) in central California. Sci. Hortic. 227, 261–271. doi: 10.1016/j.scienta.2017.09.048
Brillante, L., Martínez-Luscher, J., Yu, R., Plank, C. M., Sanchez, L., Bates, T. L., et al. (2017). Assessing spatial variability of grape skin flavonoids at the vineyard scale based on plant water status mapping. J. Agric. Food Chem. 65, 5255–5265. doi: 10.1021/acs.jafc.7b01749
Brillante, L., Mathieu, O., Lévêque, J., and Bois, B. (2016b). Ecophysiological modeling of grapevine water stress in burgundy terroirs by a machine-learning approach. Front. Plant Sci. 7:796. doi: 10.3389/fpls.2016.00796
Brillante, L., Mathieu, O., Lévêque, J., van Leeuwen, C., and Bois, B. (2018b). Water status and must composition in grapevine cv. Chardonnay with different soils and topography and a mini meta-analysis of the δ13C/water potentials correlation. J. Sci. Food Agric. 98, 691–697. doi: 10.1002/jsfa.8516
Brook, A., De Micco, V., Battipaglia, G., Erbaggio, A., Ludeno, G., Catapano, I., et al. (2020). A smart multiple spatial and temporal resolution system to support precision agriculture from satellite images: proof of concept on Aglianico vineyard. Remote Sens. Environ. 240:111679. doi: 10.1016/j.rse.2020.111679
Bustan, A., Dag, A., Yermiyahu, U., Erel, R., Presnov, E., Agam, N., et al. (2016). Fruit load governs transpiration of olive trees. Tree Physiol. 36, 380–391. doi: 10.1093/treephys/tpv138
Castellarin, S. D., Matthews, M. A., Di Gaspero, G., and Gambetta, G. A. (2007). Water deficits accelerate ripening and induce changes in gene expression regulating flavonoid biosynthesis in grape berries. Planta 227, 101–112. doi: 10.1007/s00425-007-0598-8
Cernusak, L. A., Ubierna, N., Winter, K., Holtum, J. A. M., Marshall, J. D., and Farquhar, G. D. (2013). Environmental and physiological determinants of carbon isotope discrimination in terrestrial plants. New Phytol. 200, 950–965. doi: 10.1111/nph.12423
Cernusak, L. A., Winter, K., Aranda, J., Turner, B. L., and Marshall, J. D. (2007). Transpiration efficiency of a tropical pioneer tree (Ficus insipida) in relation to soil fertility. J. Exp. Bot. 58, 3549–3566. doi: 10.1093/jxb/erm201
Chaves, M. M., Zarrouk, O., Francisco, R., Costa, J. M., Santos, T., Regalado, A. P., et al. (2010). Grapevine under deficit irrigation: hints from physiological and molecular data. Ann. Bot. 105, 661–676. doi: 10.1093/aob/mcq030
Conrad, O., Bechtel, B., Bock, M., Dietrich, H., Fischer, E., Gerlitz, L., et al. (2015). System for Automated Geoscientific Analyses (SAGA) v. 2.1.4. Geosci. Model Dev. 8, 1991–2007. doi: 10.5194/gmd-8-1991-2015
R Core Team (2018). R:A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Costantini, E. A. C., Pellegrini, S., Bucelli, P., Barbetti, R., Campagnolo, S., Storchi, P., et al. (2010). Mapping suitability for Sangiovese wine by means of δ13C and geophysical sensors in soils with moderate salinity. Eur. J. Agron. 33, 208–217. doi: 10.1016/j.eja.2010.05.007
de Souza, C. R., Maroco, J. P., dos Santos, T. P., Rodrigues, M. L., Lopes, C. M., Pereira, J. S., et al. (2005). Impact of deficit irrigation on water use efficiency and carbon isotope composition (delta13C) of field-grown grapevines under Mediterranean climate. J. Exp. Bot. 56, 2163–2172. doi: 10.1093/jxb/eri216
Diffenbaugh, N. S., and Scherer, M. (2013). Using climate impacts indicators to evaluate climate model ensembles: temperature suitability of premium winegrape cultivation in the United States. Clim. Dyn. 40, 709–729. doi: 10.1007/s00382-012-1377-1
Dutilleul, P. (1993). Modifying the t test for assessing the correlation between two spatial processes. Biometrics 49, 305–314.
Farquhar, G. D., Buckley, T. N., and Miller, J. M. (2002). Optimal stomatal control in relation to leaf area and Nitrogen content. Silva Fenn. 36, 625–637.
Farquhar, G. D., Ehleringer, J. R., and Hubick, K. T. (1989). Carbon Isotope Discrimination and Photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40, 503–537. doi: 10.1146/annurev.pp.40.060189.002443
Flexas, J., Galmés, J., Gallé, A., Gulias, J., Pou, A., Ribas-Carbo, M., et al. (2010). Improving water use efficiency in grapevines: potential physiological targets for biotechnological improvement. Aust. J. Grape Wine Res. 16, 106–121. doi: 10.1111/j.1755-0238.2009.00057.x
Gaudillère, J.-P., Van Leeuwen, C., and Ollat, N. (2002). Carbon isotope composition of sugars in grapevine, an integrated indicator of vineyard water status. J. Exp. Bot. 53, 757–763.
Gaudin, R., Roux, S., and Tisseyre, B. (2017). Linking the transpirable soil water content of a vineyard to predawn leaf water potential measurements. Agric. Water Manag. 182, 13–23. doi: 10.1016/j.agwat.2016.12.006
Gavlak, R. G., Horneck, D. A., and Miller, R. O. (1994). “Plant, soil and water reference methods for the western region,” Western Regional Extension Publication (WREP) Fairbanks: University of Alaska, 125.
Gómez-Alonso, S., and García-Romero, E. (2009). Effect of irrigation and variety on oxygen (δ18O) and carbon (δ13C) stable isotope composition of grapes cultivated in a warm climate. Aust. J. Grape Wine Res. 16, 283–289. doi: 10.1111/j.1755-0238.2009.00089.x
Guehl, J.-M., Fort, C., and Ferhi, A. (1995). Differential response of leaf conductance, carbon isotope discrimination and water- use efficiency to nitrogen deficiency in maritime pine and pedunculate oak plants. New Phytol. 131, 149–157.
Guilpart, N., Metay, A., and Gary, C. (2014). Grapevine bud fertility and number of berries per bunch are determined by water and nitrogen stress around flowering in the previous year. Eur. J. Agron. 54, 9–20. doi: 10.1016/j.eja.2013.11.002
Guix-Hébrard, N., Voltz, M., Trambouze, W., Garnier, F., Gaudillère, J. P., and Lagacherie, P. (2007). Influence of watertable depths on the variation of grapevine water status at the landscape scale. Eur. J. Agron. 27, 187–196. doi: 10.1016/j.eja.2007.03.005
Gutiérrez, S., Diago, M. P., Fernández-Novales, J., and Tardaguila, J. (2018). Vineyard water status assessment using on-the-go thermal imaging and machine learning. PLoS One 13:e0192037. doi: 10.1371/journal.pone.0192037
He, M., and Dijkstra, F. A. (2014). Drought effect on plant nitrogen and phosphorus?: a meta- analysis. New Phytol. 204, 924–931.
Herrero-Langreo, A., Tisseyre, B., Goutouly, J.-P. P., Scholasch, T., and Van Leeuwen, C. (2013). Mapping grapevine (Vitis vinifera L.) water status during the season using carbon isotope ratio (δ13C) as ancillary data. Am. J. Enol. Vitic. 64, 307–315. doi: 10.5344/ajev.2013.12125
Hoekstra, A. Y., and Mekonnen, M. M. (2012). The water footprint of humanity. Proc. Natl. Acad. Sci. U.S.A. 109, 3232–3237. doi: 10.1073/pnas.1109936109
Koundouras, S., Tsialtas, I. T., Zioziou, E., and Nikolaou, N. (2008). Rootstock effects on the adaptive strategies of grapevine (Vitis vinifera L. cv. Cabernet-Sauvignon) under contrasting water status: Leaf physiological and structural responses. Agric. Ecosyst. Environ. 128, 86–96. doi: 10.1016/j.agee.2008.05.006
Kustas, W. P., Agam, N., and Ortega-Farias, S. (2019). Forward to the GRAPEX special issue. Irrig. Sci 37, 221–226. doi: 10.1007/s00271-019-00633-7
Martínez-Lüscher, J., Brillante, L., and Kurtural, S. K. (2019). Flavonol profile is a reliable indicator to assess canopy architecture and the exposure of red wine grapes to solar radiation. Front. Plant Sci. 10:10. doi: 10.3389/fpls.2019.00010
Martínez-Lüscher, J., Brillante, L., Nelson, C. C. C., Al-Kereamy, A. M. A. M., Zhuang, S., and Kurtural, S. K. K. (2017). Precipitation before bud break and irrigation affect the response of grapevine ‘Zinfandel’ yields and berry skin phenolic composition to training systems. Sci. Hortic. 222, 153–161. doi: 10.1016/j.scienta.2017.05.011
Maupin, M. A., Kenny, J. F., Hutson, S. S., Lovelace, J. K., Barber, N. L., and Linsey, K. S. (2014). Estimated use of water in the United States in 2010: U.S. Geological Survey Circular 1405, 56. doi: 10.3133/cir1405
Medrano, H., Tomás, M., Martorell, S., Escalona, J.-M., Pou, A., Fuentes, S., et al. (2015). Improving water use efficiency of vineyards in semi-arid regions. Rev. Agron. Sustain. Dev. 35, 499–517. doi: 10.1007/s13593-014-0280-z
Miner, G. L., Bauerle, W. L., and Baldocchi, D. D. (2017). Estimating the sensitivity of stomatal conductance to photosynthesis: a review. Plant Cell Environ. 40, 1214–1238. doi: 10.1111/pce.12871
Myers, N., Mittermeler, R. A., Mittermeler, C. G., Da Fonseca, G. A. B., and Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Pascual, M., Villar, J. M., and Rufat, J. (2016). Water use efficiency in peach trees over a four-years experiment on the effects of irrigation and nitrogen application. Agric. Water Manag. 164, 253–266. doi: 10.1016/j.agwat.2015.10.021
Pebesma, E. J. (2004). Multivariable geostatistics in S: the gstat package. Comput. Geosci. 30, 683–691. doi: 10.1016/j.cageo.2004.03.012
Poni, S., Bernizzoni, F., Civardi, S., Gatti, M., Porro, D., and Camin, F. (2009). Performance and water-use efficiency (single-leaf vs. whole-canopy) of well-watered and half-stressed split-root Lambrusco grapevines grown in Po Valley (Italy). Agric. Ecosyst. Environ. 129, 97–106. doi: 10.1016/j.agee.2008.07.009
Ritchey, J. G., and Andrew, L. A. (1999). A standard red wine: monomeric analysis of commercial Cabernet-Sauvignon wines. Am. J. Enol. Vitic. 50, 91–100.
Sanchez, L. A., Sams, B., Alsina, M. M., Hinds, N., Klein, L. J., and Dokoozlian, N. (2017). Improving vineyard water use efficiency and yield with variable rate irrigation in California. Adv. Anim. Biosci. 8, 574–577. doi: 10.1017/s2040470017000772
Santesteban, L. G., Miranda, C., and Royo, J. B. (2016). Interest of carbon isotope ratio (δ13C) as a modeling tool of grapevine yield, berry size and sugar content at within-field, winegrowing domain and regional scale. Theor. Exp. Plant Physiol. 28, 193–203. doi: 10.1007/s40626-016-0067-5
Suter, B., Triolo, R., Pernet, D., Dai, Z., and Van Leeuwen, C. (2019). Modeling stem water potential by separating the effects of soil water availability and climatic conditions on water status in grapevine (Vitis vinifera L.). Front. Plant Sci. 10:1485. doi: 10.3389/fpls.2019.01485
Tao, Y., Zhang, Y., Jin, X., Saiz, G., Jing, R., Guo, L., et al. (2015). More rice with less water – evaluation of yield and resource use efficiency in ground cover rice production system with transplanting. Eur. J. Agron. 68, 13–21. doi: 10.1016/j.eja.2015.04.002
Taylor, J. A., Acevedo-Opazo, C., Ojeda, H., and Tisseyre, B. (2010). Identification and significance of sources of spatial variation in grapevine water status. Aust. J. Grape Wine Res. 16, 218–226. doi: 10.1111/j.1755-0238.2009.00066.x
van Leeuwen, C., Friant, P., Choné, X., Tregoat, O., Koundouras, S., and Dubourdieu, D. (2004). Influence of climate, soil, and cultivar on terroir. Am. J. Enol. Vitic. 55, 207–217.
von Caemmerer, S., Ghannoum, O., Pengelly, J. J. L., and Cousins, A. B. (2014). Carbon isotope discrimination as a tool to explore C4 photosynthesis. J. Exp. Bot. 65, 3459–3470. doi: 10.1093/jxb/eru127
Weih, M., Bonosi, L., Ghelardini, L., and Rönnberg-Wästljung, A. C. (2011). Optimizing nitrogen economy under drought: increased leaf nitrogen is an acclimation to water stress in willow (Salix spp.). Ann. Bot. 108, 1347–1353. doi: 10.1093/aob/mcr227
White, M. A., Diffenbaugh, N. S., Jones, G. V., Pal, J. S., and Giorgi, F. (2006). Extreme heat reduces and shifts United States premium wine production in the 21st century. Proc. Natl. Acad. Sci. U.S.A. 103, 11217–11222. doi: 10.1073/pnas.0603230103
Williams, L. E., and Baeza, P. (2007). Relationships among ambient temperature and vapor pressure deficit and leaf and stem water potentials of fully irrigated, field-grown grapevines. Am. J. Enol. Vitic. 58, 173–181.
Keywords: water, precision agriculture, anthocyanins, nitrogen, terroir, irrigation, zoning, selective harvest
Citation: Brillante L, Martínez-Lüscher J, Yu R and Kurtural SK (2020) Carbon Isotope Discrimination (δ13 C) of Grape Musts Is a Reliable Tool for Zoning and the Physiological Ground-Truthing of Sensor Maps in Precision Viticulture. Front. Environ. Sci. 8:561477. doi: 10.3389/fenvs.2020.561477
Received: 12 May 2020; Accepted: 04 September 2020;
Published: 24 September 2020.
Edited by:
Sandro Conticelli, University of Florence, ItalyReviewed by:
Gianluca Bianchini, University of Ferrara, ItalyNives Ogrinc, Institut Jožef Stefan (IJS), Slovenia
Copyright © 2020 Brillante, Martínez-Lüscher, Yu and Kurtural. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luca Brillante, bHVjYWJyaWxsYW50ZUBjc3VmcmVzbm8uZWR1