
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Endocrinol. , 28 June 2019
Sec. Cancer Endocrinology
Volume 10 - 2019 | https://doi.org/10.3389/fendo.2019.00417
This article is part of the Research Topic Testicular Cancer: New Insights on the Origin, Genetics, Treatment, Fertility, General Health, Quality of Life and Sexual Function View all 18 articles
 Murilo Bonatelli1
Murilo Bonatelli1 Eduardo C. A. Silva2
Eduardo C. A. Silva2 Flavio M. Cárcano3,4Maurício G. Zaia1Luiz F. Lopes5
Flavio M. Cárcano3,4Maurício G. Zaia1Luiz F. Lopes5 Cristovam Scapulatempo-Neto1,2
Cristovam Scapulatempo-Neto1,2 Céline Pinheiro1,4*
Céline Pinheiro1,4*Testicular Germ Cell Tumors (TGCTs) are a rare group of neoplasms and the most common solid malignancy arising in young male adults. Despite the good response of these tumors to platinum-based chemotherapy, some patients are refractory to treatment and present poor clinical outcomes. During carcinogenesis and tumor development, cancer cells reprogram energy metabolism toward a hyper-glycolytic phenotype, an emerging hallmark of cancer. This phenomenon, known as the Warburg effect or aerobic glycolysis, involves overexpression of metabolism-related proteins, like glucose and monocarboxylate transporters, pH regulators and intracellular glycolytic enzymes. The metabolic profile of TGCTs is very little explored and, recently, this metabolic rewiring of cancer cells has been associated with aggressive clinicopathological characteristics of these tumors. The overexpression of monocarboxylate transporter 4 (MCT4) in TGCTs has been pointed out as a poor prognostic factor, as well as a promising therapeutic target. As a result, the main aim of the present study was to evaluate the prognostic value of key metabolism-related proteins in TGCTs. The immunohistochemical expressions of CD44 (as a monocarboxylate transporter chaperone), glucose transporter 1 (GLUT1), carbonic anhydrase IX (CAIX), hexokinase II (HKII) and lactate dehydrogenase V (LDHV) were evaluated in a series of 148 adult male patients with TGCTs and associated with clinicopathological parameters. In addition, paired normal tissues were also evaluated. The sample included 75 seminoma and 73 non-seminoma tumors. GLUT1 and CD44 expression was significantly increased in malignant samples when compared to paired normal samples. Conversely, HKII and LDHV expressions were significantly decreased in malignant samples. Concerning the clinicopathological values, CAIX expression was significantly associated with disease recurrence, while HKII expression was significantly associated with aggressive characteristics of TGCTs, including higher staging and non-seminoma histology. In conclusion, this study brings new insights on the metabolic characteristics of TGCTs, showing alterations in the expression of proteins related with the Warburg effect, as well as associations of the hyper-glycolytic and acid-resistant phenotype with aggressive clinicopathological parameters.
Testicular germ cell tumors (TGCTs) are the most frequent solid malignancies arising in young male adults (1, 2) and show an increase in incidence throughout the last decades, especially in Europea-descendent men (1, 3, 4). Divided into two major histological types, homogeneous seminoma and heterogeneous non-seminoma tumors (1), TGCTs tend to have a good response to platinum-based chemotherapy, with seminomas presenting more favorable outcomes in comparison to non-seminoma tumors (2). Despite the high rates of cure—over 90% in patients with early diagnosed disease (independent of histological type) (1, 5)—about 10–20% are refractory to treatment and present unfavorable clinical outcomes (6, 7).
The major mechanisms involved in the development of TGCTs are copy number variations (aneuploidies) and some recurrent somatic mutations. The isochromosome 12p is present in almost all tumors and is considered a marker for TGCTs. In fact, the underlying mechanisms involving isochromosome 12p in the development of TGCTs are still unclear, but there is enough evidence that implicate this alteration as an earlier triggering event, leading to invasiveness growth and malignization. Furthermore, the most frequently mutated driver oncogenes found in seminomas are KIT and KRAS, with 25–30 and 5–10% mutation frequencies, respectively (2, 4). Besides that, there is still a lack of information in understanding the complex heterogeneity of TGCTs, which highlights the importance of the discovery of different oncogenic events involving these tumors to optimize treatment and management. In this context, the recently described hallmark of cancer of deregulation of cellular energetics is gaining additional attention in the last years and should be considered as a possible relevant biological mechanism in TGCTs (8, 9).
During carcinogenesis and tumor development, cancer cells reprogram energy metabolism toward a hyper-glycolytic phenotype, even in the presence of high oxygen levels. This phenomenon, known as the Warburg effect or aerobic glycolysis, leads to a higher production of lactate than the normal metabolic phenotype, which relies mostly on oxidative phosphorylation (10–12). To fuel all the energy required and avoid intracellular acidification and apoptosis, cancer cells upregulate some key proteins, like glucose and monocarboxylate transporters, pH regulators and intracellular glycolytic enzymes (13). In comparison to oxidative phosphorylation, glycolysis is not an energetic efficient pathway but is a faster way to provide energy, metabolic intermediates, and biochemical building blocks, essential for anabolic reactions, and thus enhancing the aggressive characteristics presented by malignant cells (14). As demonstrated by many studies (15, 16), the overexpression of metabolism-related proteins plays an important role in the development and maintenance of the malignant phenotype of a vast majority of tumors. In this context, these metabolic players have been pointed out as prognostic factors and can be explored as promising therapeutic targets.
Although studies evaluating the implication of metabolic rewiring in the development and progression of TGCTs are lacking, there is evidence that these tumors present a highly glycolytic behavior, mainly attributed to their elevated levels of glucose consumption as demonstrated by 18F-fluorodeoxyglucose positron emission tomography (FDG-PET) studies (17–20). Furthermore, when comparing malignant and benign samples, a recent study (21) shows that TGCTs overexpress monocarboxylate transporter 4 (MCT4) and its chaperone CD147. Additionally, the upregulation of monocarboxylate transporter 1 (MCT1), MCT4 and CD147 has been associated with aggressive clinicopathological characteristics of these tumors, while MCT4 overexpression was associated with a worse prognosis, with patients presenting a shorter overall and event-free survival. Other studies show that TGCTs have an increase in the expression of glucose transporter 3 (GLUT3) protein (22), which is often found overexpressed by malignant neoplasms (23, 24).
Since metabolic reprogramming in human tumors need to be further exploited, the study of different metabolism-related proteins may offer a better understanding about their role, relevance, and implication in the biological characteristics and the complex heterogeneity of TGCTs. In this context, CD44, a chaperone for proper localization and activity of MCT1 and MCT4 in the plasma membrane, glucose transporter 1 (GLUT1), the pH regulator carbonic anhydrase 9 (CAIX), as well as hexokinase II (HKII), responsible for the irreversible glucose phosphorylation in the earlier steps of glycolysis and lactate dehydrogenase V (LDHV), the isoenzyme with higher affinity for pyruvate, that catalyze the conversion of pyruvate into lactate, arise as key players in the metabolic reprogramming of cancer cells (13, 16, 25, 26).
Therefore, regarding the emerging role of metabolic rewiring in tumors, as well as the biological complexity and the absence of studies considering this context in TGCTs, the aims of this study were to evaluate the expression of CD44, GLUT1, CAIX, HKII, and LDHV in TGCTs and normal samples, using tissue microarrays (TMAs), and to associate the expression with clinicopathological data to determine whether these proteins have some biological and/or prognostic value.
The series included 148 formalin-fixed paraffin embedded adult TGCTs samples, retrieved from the Pathology Department of Barretos Cancer Hospital, from 2007 to 2013. Only primary tumors, prior to chemotherapy, were selected. Additionally, paired normal samples were collected and analyzed when available (n = 66 for CD44, n = 59 for GLUT1, n = 78 for CAIX, n = 87 for HKII and n = 84 for LDHV; the different number of normal samples analyzed for each protein is related to sample loss as a result of block sectioning). The clinicopathological data included age, date of diagnosis, histological types, grading, staging (TNM), presence of vascular invasion, International Germ Cell Cancer Collaborative Group (IGCCCG) stratification risk (63), and dates of surgery, chemotherapy, recurrence, progression and death. Patients' mean age was 32.3 years (ranging from 18 to 73 years) and most of them were caucasian (62.2%). Detailed information on the clinicopathological data of the sample is depicted in Table 1. This study was approved by the Ethics Committee on Research of Barretos Cancer Hospital (number 541235).
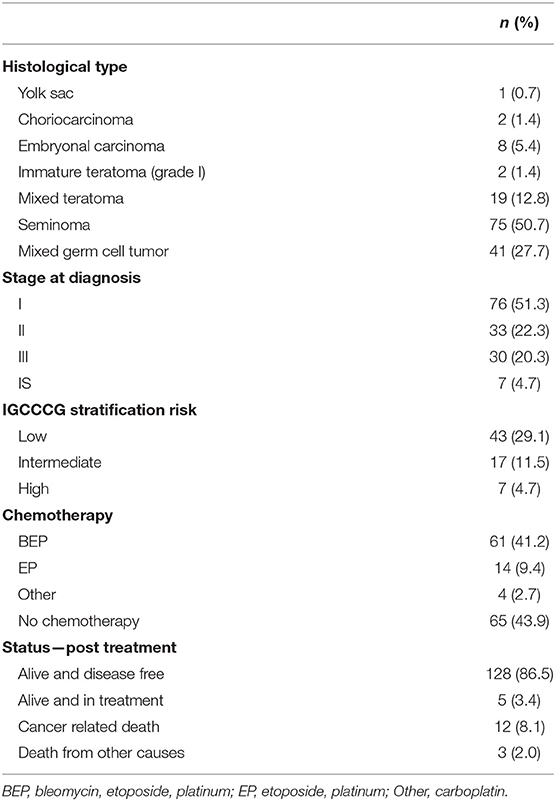
Table 1. Clinicopathological characteristics of adult testicular germ cell tumor patients.
TMAs were constructed for the immunohistochemical reactions. All the cases were reviewed by an experienced pathologist (ECAS) for diagnostic confirmation and demarcation of tumor areas for TMA cores. Each TMA contained sample cores of 1.0 mm diameter from all histological subtypes and corresponding normal tissues, in triplicate. Liver, kidney and placenta were used as controls for TMA orientation.
Immunohistochemistry for GLUT1 and CAIX was performed using a streptavidin-biotin-peroxidase complex (Ultravision Detection System: Large Volume Anti-Polyvalent, HRP, Lab Vision Corporation, Fremont, CA), according to manufacturer's instructions and as previously described (27). Immunohistochemistry for CD44 was performed using a biotin-free principle (ADVANCE HRP, Dako, Carpinteria, CA), according to manufacturer's instructions. For HKII and LDHV, the reactions were performed using an avidin-biotin-peroxidase complex principle (R.T.U. VECTASTAIN Kit, Vector Laboratories, Burlingame, CA), according to manufacturer's instructions. Details on antigen retrieval and each antibody used are described in Table 2. For visualization, slides were incubated with 3,3′-diamino-benzidine (Liquid DAB+ Substrate Chromogen System, Dako, Carpinteria, CA), according to manufacturer's instructions, then counterstained with hematoxylin and permanently mounted. As positive controls, placenta was used for GLUT1, normal gastric mucosa for CAIX and squamous cell carcinoma of oral cavity for CD44, HKII, and LDHV. Negative controls were available in the same tissue sections used as positive controls.

Table 2. Detailed aspects of immunohistochemistry.
TMAs and whole sections were scored semi-quantitatively for extension of expression in cancer cells as follows: 0: no immunoreactive cells; 1: <5% of immunoreactive cells; 2: 5–50% of immunoreactive cells; and 3: >50% of immunoreactive cells. Also, intensity of staining was scored semi-qualitatively as follows: 0: negative; 1: weak; 2: intermediate; and 3: strong. The final score was defined as the sum of both parameters (extension and intensity) and grouped as negative (score 0–2) and positive (score 3–6), as previously described (28, 29). Only protein expression in plasma membrane was considered for CD44, GLUT1, and CAIX analysis, while for HKII and LDHV only cytoplasmic expression was considered for further analysis. TMAs were evaluated by two experienced pathologists independently (ECAS and CS-N). Discordant cases were reviewed and scored in consensus.
Data collected was analyzed using IBM SPSS Statistics software (version 23.0, IBM Company, Armonk, NY). During immunohistochemical evaluation, loss of tumor representativity in TMA cores as well as whole core loss was observed, influencing the final number of cases used for statistical analysis. Frequency of protein expression in normal and malignant tissues was compared using McNemar's test, while comparison with clinicopathological data was analyzed using Pearson's chi-square test and Fisher's exact test, according to the sample's characteristics. Overall survival was defined as the time from the date of primary diagnosis to last follow-up or death. Event-free survival was defined as the time from the primary diagnosis to the event date (recurrence, disease progression or death). None of the patients presented secondary tumors until the last follow-up. For survival models, only stage II and III patients were considered for further analysis. Overall and event-free survival curves were constructed using Kaplan-Meier's method and the data compared with log-rank test. Multivariate analysis by Cox proportional hazards regression model was used to determine independent predictors of survival. Independent variables were analyzed by univariate analysis, followed by multivariate analysis of all variables that reached a p < 0.2 at univariate analysis. For all tests, the level of significance established was 5% (significant results if p < 0.05).
Immuhistochemical evaluation of adult testicular germ cell tumors showed that expression of CD44, GLUT1 and CAIX was mostly exclusively found in plasma membrane. Regarding the expression of HKII and LDHV in tumor samples, both proteins were mostly detected in cytoplasm. According to the results observed in tumors, paired normal tissues showed similar expression patterns, with CD44, GLUT1 and CAIX frequently found in plasma membrane and HKII and LDHV in cytoplasm (Figure 1).
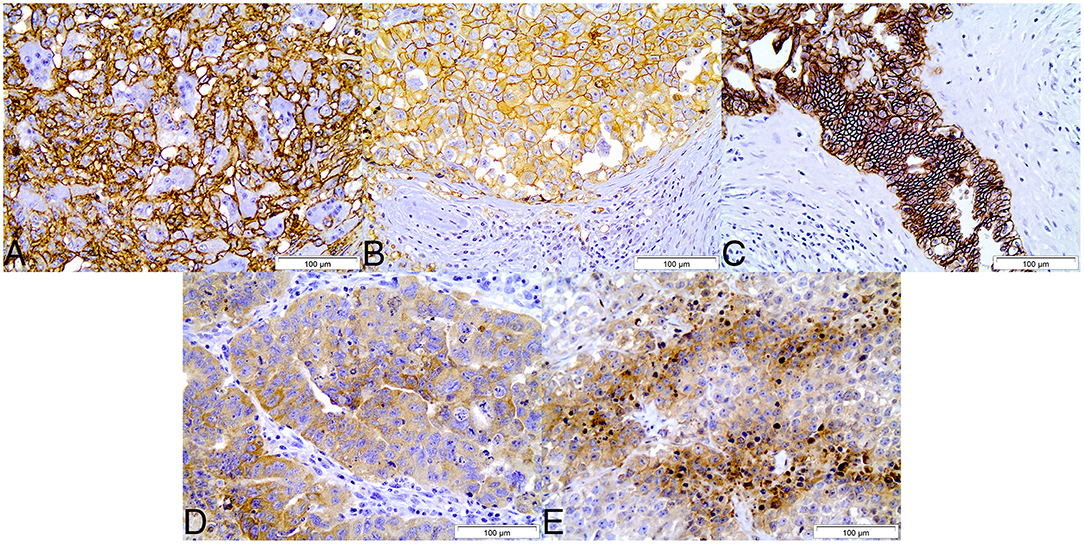
Figure 1. Immunohistochemical expression of CD44 (A), GLUT1 (B), CAIX (C), HKII (D), and LDHV (E) in adult testicular germ cell tumors. CD44, GLUT1, and CAIX show plasma membrane staining, while HKII and LDHV show cytoplasmic staining. Magnification: 400x.
Comparison of protein expression between tumor samples and paired normal tissues showed a significantly increased expression of CD44 and GLUT1 in tumor samples (p = 0.004 and p < 0.001, Figure 2). Conversely, HKII and LDHV expression was significantly decreased in malignant samples (p < 0.001 and p < 0.001, Figure 2).
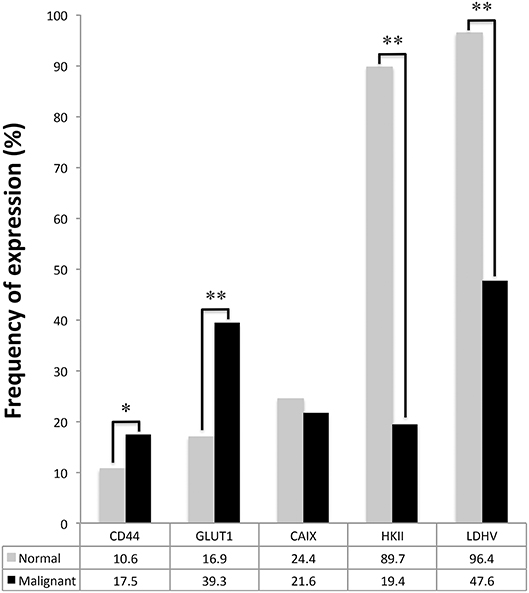
Figure 2. Plasma membrane expression of GLUT1, CAIX, and CD44 and cytoplasmic expression of HKII and LDHV in adult testicular germ cell tumors and correspondent normal testicular tissues. McNemar's test was used to assess differences of expression frequency between tumor and normal tissue. *p = 0.004, **p < 0.001.
The associations between the expression of the metabolism-related proteins and the clinicopathological data of the patients are shown in Table 3. CAIX expression presented a significant association with recurrence (p = 0.024), while HKII showed a statistically significant association with non-seminoma tumors (p < 0.001) and higher stages (p = 0.019). CD44, GLUT1 and LDHV showed no significant associations with clinicopathological parameters.
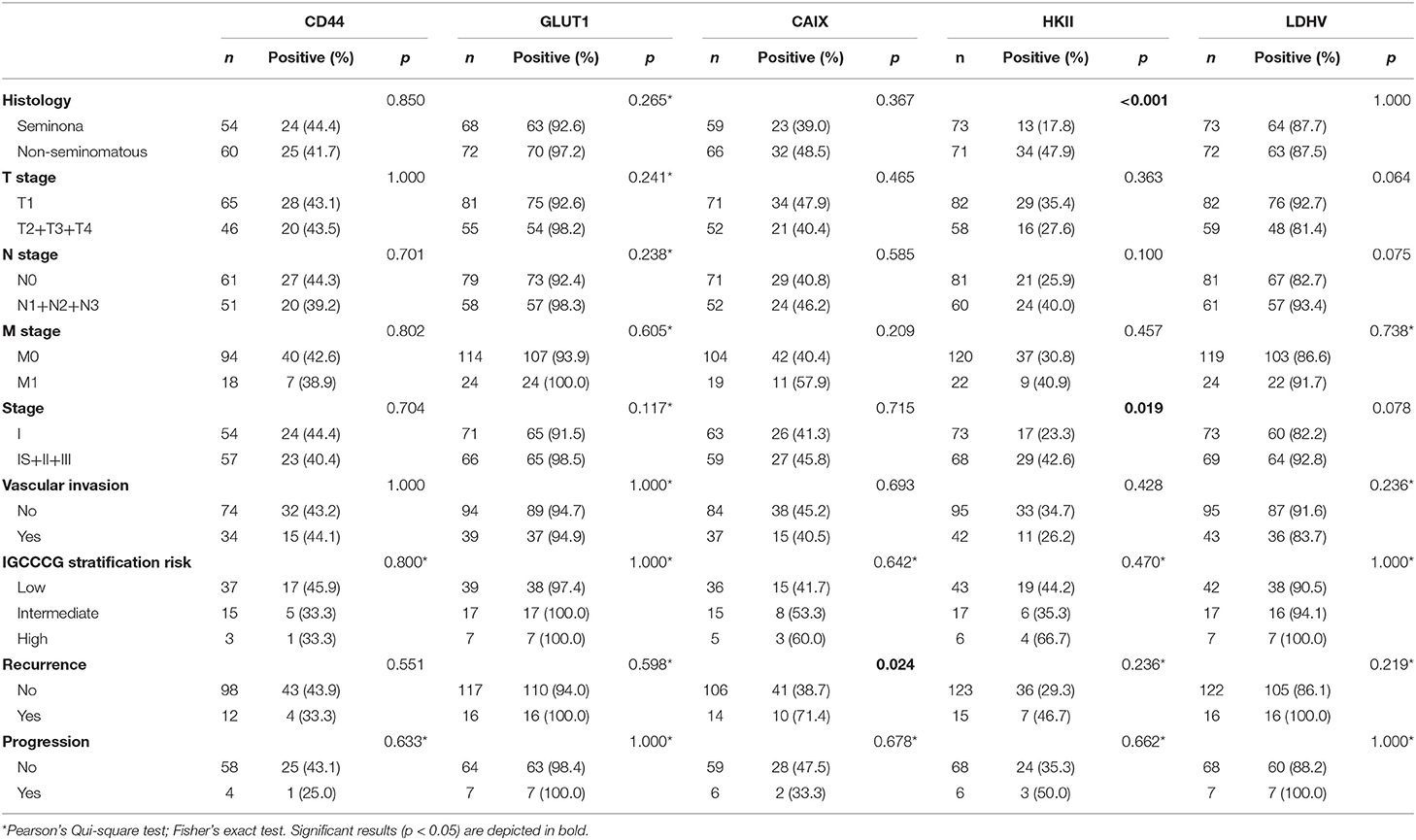
Table 3. Association of CD44, GLUT1 and CAIX membrane expression and cytoplasmic expression of HKII and LDHV with clinicopathological characteristics of adult testicular germ cell tumors.
Kaplan-Meier analysis for the expression of metabolism-related proteins showed no significant associations with overall and event-free survival (data not shown). The predictive prognostic values of the proteins and clinicopathological parameters for overall survival and event-free survival were analyzed by means of Cox proportional hazards regression models (Tables 4, 5, respectively). Univariate analysis revealed predictive prognostic values for stage III and IGCCCG intermediate and high risk stratifications in overall and event-free survival. Multivariate analysis showed that IGCCCG high risk classification was an independent predictor for overall and event-free survival (HR: 9.987 p = 0.034 and HR: 11.061 p = 0.014, respectively). None of the analyzed proteins presented a prognostic value in TGCTs patients.
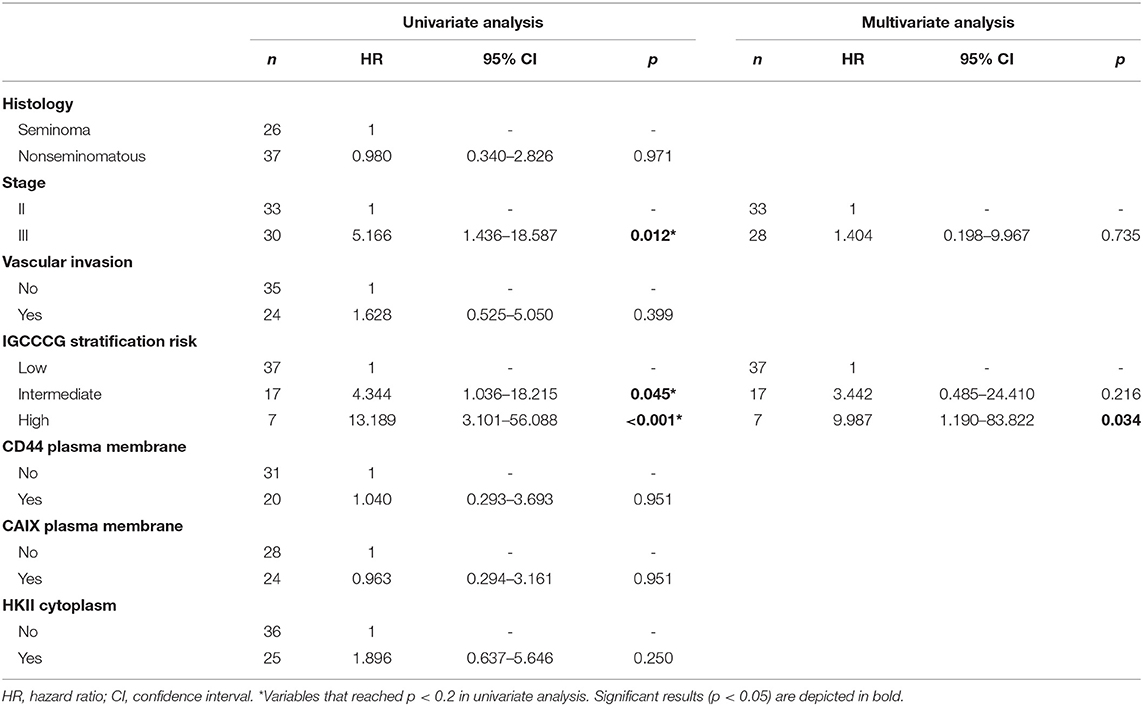
Table 4. Prognostic factors for overall survival in adult testicular germ cell tumors.
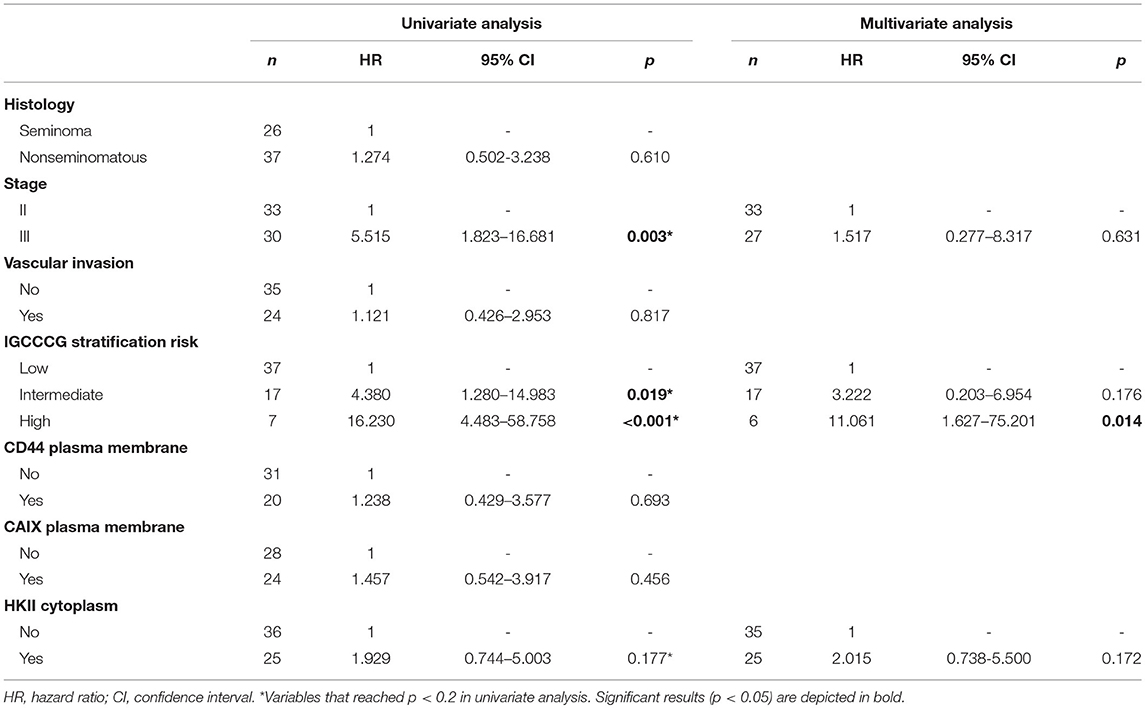
Table 5. Prognostic factors for event-free survival in adult testicular germ cell tumors.
Our study showed an increase in the expression frequency of GLUT1 and CD44 in adult testicular germ cell tumors. In a previous study, the authors showed that MCT4 and CD147 expression was increased in TGCTs patients (21). Taken together, these results suggest a metabolic reprogramming of malignant cells toward a hyperglycolytic and acid-resistant phenotype in TGCTs.
The high frequency of GLUT1 expression is in agreement with the natural behavior of cancer cells as high glucose consumers. In fact, glucose transporters need to be overexpressed to fuel part of the metabolic reprogramming required by malignant cells to produce energy and intermediates for anabolism, as well as regulate redox state (30). This metabolic reprogramming demands an increased uptake of glucose, mainly provided by GLUT1 (31). In addition, CD44 expression is related to the overexpression of MCT4 previously described in TGCTs (21), suggesting that CD44 and MCT4 work together in lactate efflux (25), favoring the Warburg effect.
Regarding the association of metabolism-related proteins expression with clinicopathological characteristics of TGCTs, our study showed a significant association between CAIX status and recurrence, as well as HKII positive expression with aggressive clinicopathological parameters (non-seminoma tumors and higher staging). CAIX, a hypoxia marker, exerts a pH control regulation, which contributes to the acid-mediated cancer cell invasiveness when overexpressed (32–35), and has been associated with a worse prognosis in a variety of tumors (34). Also, the relation of CAIX expression with recurrence in TGCTs corroborates the role of CAIX in stimulating the invasion and aggressive phenotype of malignant cells and this association had been demonstrated by two studies (36, 37). In the firsty study (36), the authors show that CAIX serum levels was higher in TGCTs metastatic patients when compared to healthy controls, and that CAIX serum levels are significantly associated with intratumoral CAIX expression. However, CAIX serum levels do not demonstrate association with clinicopathlogical data, neither a prognostic value in clinical outcomes. According to the second study (37), the authors show a significant increase in the expression of CAIX in TGCTs samples compared to paired adjacent normal samples. This result, not reached by our study, probably can be explained by the numerical sample difference between the two works. Moreover, the authors show that CAIX positive expression was significantly associated with a worse progression-free survival, predominatly founded in patients with metastatic disease, which is in agreement with the association of CAIX positive expression with recurrence, demonstrated in the present study. Furthermore, the significant association of HKII positive expression with non-seminoma tumors and higher stages demonstrated by our study is in accordance with the role of this protein in providing energy for tumors, leading to disease progression and poor outcomes. Regardless of the lack of information about HKII expression in TGCTs patients, several studies associated HKII overexpression with aggressive characteristics and worse outcomes in different types of neoplasms, such as breast (38–40), cervical (41), colorectal (42), glioblastoma (43), liver (44, 45), lung (39), pancreatic (46, 47) and prostate (48). Indeed, HKII is described as one of the main proteins responsible for mediating the Warburg effect in cancer cells. This protein binds to the voltage dependent anion channel (VDAC) in mitochondria outer membrane, keeping the channel in an open state, gaining direct access to ATP generated intra-mitochondrially for glucose phosphorylation (49). This is in accordance with in vitro studies demonstrating the role of VDAC-bound HKII in supporting the Warburg effect (50). Additionally, other activities have been attributed to HKII in cancer metabolism context, which favors the aggressive phenotype of malignant cells, including the production of antioxidant molecules, direct protection of mitochondria against redox stress (anti-apoptotic effect) and facilitation of autophagy under starvation. Moreover, HIF-1α stimulation by AKT and mTORC1 has been described as the mechanism mainly responsible for HKII upregulation (51).
In contrast to GLUT1 and CD44 expression, a decreased frequency of HKII and LDHV expression in tumor samples, compared to normal testicular samples, was observed in the present study. Due to insufficient information available in the literature about the expression of HKII in normal testis, a search in the Human Protein Atlas (HPA) database, which integrates the protein expression profiles of 44 normal human tissues to RNA sequencing data of 32 out of these 44 tissue types (52), was conducted. HPA data showed that HKII RNA and protein expressions were correlated and more pronounced in male normal tissues, like testis and epididymis, corroborating our results. Also, a study done by Postic and collaborators (53), showed a relation between hexokinases and glucose transporters isoforms. Using rat models, the authors demonstrated that, during embryonic development, HKI and GLUT1 were the major isoforms expressed and related to energy production. After weaning, with the acquisition of insulin-sensitivity tissues, there is a switch in both isoforms, with HKII and GLUT4 participating mostly in energy production. These results suggest the important role of HKII in normal tissues after embryonic development for energy production. Our results are in accordance with this previous data as malignant testicular tissues tend to resemble the hypoxic and undifferentiated embryonic tissues (54, 55), therefore showing higher expression of GLUT1 but lower expression of HKII when compared to normal tissues. Regarding LDHV expression, Dodo and collaborators (56) found that LDHV was co-expressed with LDHX, the major LDH isoform present in testis, which has been found in different animals, including humans (57, 58). Curiously, LDHX mice knock-out presented an ablation of LDHV expression, not demonstrated by wild type mice, resulting in reduced energy production through glycolysis and impaired fertilization (56). Also, another study described that LDHV human transgene expression was able to restore LDHX expression in testis and sperm of LDHX knock-out mice, also restoring sperm motility and fertilization capacity (59). Taken together, these results suggest an important role of LDHV in aerobic glycolysis presented by normal testis, indicating that this protein is required to establish proper physiologic conditions for fertilization.
The natural behavior of cancer cells in reprogramming their metabolism, with a heavier reliance in aerobic glycolysis, provides a solid field for the development of anticancer therapies. Across the decades, different glycolytic inhibitors have been tested in pre-clinical studies and clinical trials, trying to kill cancer cells by pharmacological inhibition of glucose consumption and achieve therapeutic selectivity. Pelicano and co-workers (60) discuss about the use of three potent hexokinase inhibitors: 3-bromopiruvate (3-BP), 2-deoxyglucose (2-DG), and lonidamine. The major inhibitory mechanism of these compounds was the blockage of glucose phosphorylation, mediating the uncoupling of HK from mitochondria, leading to a rapid depletion of cellular ATP. Additionally, in vitro studies show a relevant therapeutic effect of the anti-hyperglycemic drug metformin in glycolytic addicted tumors, through the inhibition of HK function (38, 61). Finally, a recent study showed the efficacy of the antifungal drugs—ketoconazole and posaconazole—in glioblastoma cells, with selective inhibition of HKII. Using in vitro and in vivo models, the authors showed that both drugs were able to increase survival of mice and decrease cell proliferation and tumor metabolism (62). Importantly, both drugs are enrolled in an early phase I clinical trial in high grade gliomas (clinicaltrials.gov—NCT03763396). Despite the promising efficacy of the use of glycolytic inhibitors for glucose addicted tumors, TGCTs show a more complex biology. Our results demonstrate that HKII and LDHV have a role in normal testis, suggesting the importance of these markers in fertilization. Indeed, the use of HK inhibitors in TGCTs patients may lead to an increase in cytotoxicity and even infertility.
The present study, together with the study done by Silva and co-workers (21), corroborates that TGCTs present a switch in cellular metabolism toward a hyper glycolytic and acid-resistant phenotype, mainly associated with a worse prognosis. Additionally, HKII appears as a marker of tumor aggressiveness, bringing new insights about the metabolic characteristics of these tumors. Although our results showed a bone fide characterization of TGCTs metabolism, further studies are warranted to achieve a better understand, especially about HKII role in testicular malignancies, providing new evidences for future therapeutic strategies.
The datasets generated for this study are available on request to the corresponding author.
This study was approved by the Ethics Committee on Research of Barretos Cancer Hospital (number 541235). Written informed consent was waived as this study was considered a minimal-risk study.
MB performed immunohistochemical reactions, statistical analysis, and wrote the manuscript. MZ performed immunohistochemical reactions. ES and CS-N analyzed histological sections and performed the immunohistochemical evaluations. FC and LL aided in the study design and discussion of the results. CP was responsible for the study design, contributed to the discussion of the results, organization, and review of the manuscript. All authors read and approved the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This study was supported by CNPq, Conselho Nacional de Desenvolvimento Científico e Tecnológico, Brasil (480119/2013-9 and 306195/2016-0).
1. Rajpert-De Meyts E, McGlynn KA, Okamoto K, Jewett MA, Bokemeyer C. Testicular germ cell tumours. Lancet. (2016) 387:1762–74. doi: 10.1016/S0140-6736(15)00991-5
2. Shen H, Shih J, Hollern DP, Wang L, Bowlby R, Tickoo SK, et al. Integrated molecular characterization of testicular germ cell tumors. Cell Rep. (2018) 23:3392–406. doi: 10.1016/j.celrep.2018.05.039
3. Horwich A, Nicol D, Huddart R. Testicular germ cell tumours. BMJ. (2013) 347:f5526. doi: 10.1136/bmj.f5526
4. Litchfield K, Levy M, Huddart RA, Shipley J, Turnbull C. The genomic landscape of testicular germ cell tumours: from susceptibility to treatment. Nat Rev Urol. (2016) 13:409–19. doi: 10.1038/nrurol.2016.107
5. Hanna N, Einhorn LH. Testicular cancer: a reflection on 50 years of discovery. J Clin Oncol. (2014) 32:3085–92. doi: 10.1200/JCO.2014.56.0896
6. Feldman DR, Bosl GJ, Sheinfeld J, Motzer RJ. Medical treatment of advanced testicular cancer. JAMA. (2008) 299:672–84. doi: 10.1001/jama.299.6.672
7. Piulats JM, Jimenez L, Garcia del Muro X, Villanueva A, Vinals F, Germa-Lluch JR. Molecular mechanisms behind the resistance of cisplatin in germ cell tumours. Clin Transl Oncol. (2009) 11:780–6. doi: 10.1007/s12094-009-0446-3
8. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. (2011) 144:646–74. doi: 10.1016/j.cell.2011.02.013
9. Pavlova NN, Thompson CB. The emerging hallmarks of cancer metabolism. Cell Metab. (2016) 23:27–47. doi: 10.1016/j.cmet.2015.12.006
10. Warburg O, Wind F, Negelein E. The metabolism of tumors in the body. J Gen Physiol. (1927) 8:519–30.
12. Koppenol WH, Bounds PL, Dang CV. Otto Warburg's contributions to current concepts of cancer metabolism. Nat Rev Cancer. (2011) 11:325–37. doi: 10.1038/nrc3038
13. Hay N. Reprogramming glucose metabolism in cancer: can it be exploited for cancer therapy? Nat Rev Cancer. (2016) 16:635–49. doi: 10.1038/nrc.2016.77
14. Deberardinis RJ, Sayed N, Ditsworth D, Thompson CB. Brick by brick: metabolism and tumor cell growth. Curr Opin Genet Dev. (2008) 18:54–61. doi: 10.1016/j.gde.2008.02.003
15. Pinheiro C, Longatto-Filho A, Azevedo-Silva J, Casal M, Schmitt FC, Baltazar F. Role of monocarboxylate transporters in human cancers: state of the art. J Bioenerg Biomembr. (2012) 44:127–39. doi: 10.1007/s10863-012-9428-1
16. Granja S, Pinheiro C, Reis RM, Martinho O, Baltazar F. Glucose addiction in cancer therapy: advances and drawbacks. Curr Drug Metab. (2015) 16:221–42. doi: 10.2174/1389200216666150602145145
17. Cremerius U, Wildberger JE, Borchers H, Zimny M, Jakse G, Gunther RW, et al. Does positron emission tomography using 18-fluoro-2-deoxyglucose improve clinical staging of testicular cancer?–Results of a study in 50 patients. Urology. (1999) 54:900–4.
18. Oechsle K, Hartmann M, Brenner W, Venz S, Weissbach L, Franzius C, et al. [18F]Fluorodeoxyglucose positron emission tomography in nonseminomatous germ cell tumors after chemotherapy: the German multicenter positron emission tomography study group. J Clin Oncol. (2008) 26:5930–5. doi: 10.1200/JCO.2008.17.1157
19. Bachner M, Loriot Y, Gross-Goupil M, Zucali PA, Horwich A, Germa-Lluch JR, et al. 2-8fluoro-deoxy-D-glucose positron emission tomography (FDG-PET) for postchemotherapy seminoma residual lesions: a retrospective validation of the SEMPET trial. Ann Oncol. (2012) 23:59–64. doi: 10.1093/annonc/mdr052
20. Dotzauer R, Thomas C, Jager W. The use of F-FDG PET/CT in testicular cancer. Transl Androl Urol. (2018) 7:875–8. doi: 10.21037/tau.2018.09.08
21. Silva ECA, Carcano FM, Bonatelli M, Zaia MG, Morais-Santos F, Baltazar F, et al. The clinicopathological significance of monocarboxylate transporters in testicular germ cell tumors. Oncotarget. (2018) 9:20386–98. doi: 10.18632/oncotarget.24910
22. Howitt BE, Brooks JD, Jones S, Higgins JP. Identification and characterization of 2 testicular germ cell markers, Glut3 and CyclinA2. Appl Immunohistochem Mol Morphol. (2013) 21:401–7. doi: 10.1097/PAI.0b013e31827b505f
23. Younes M, Lechago LV, Somoano JR, Mosharaf M, Lechago J. Immunohistochemical detection of Glut3 in human tumors and normal tissues. Anticancer Res. (1997) 17:2747–50.
24. Vander Heiden MG. Targeting cancer metabolism: a therapeutic window opens. Nat Rev Drug Discov. (2011) 10:671–84. doi: 10.1038/nrd3504
25. Slomiany MG, Grass GD, Robertson AD, Yang XY, Maria BL, Beeson C, et al. Hyaluronan, CD44, and emmprin regulate lactate efflux and membrane localization of monocarboxylate transporters in human breast carcinoma cells. Cancer Res. (2009) 69:1293–301. doi: 10.1158/0008-5472.CAN-08-2491
26. Pinheiro C, Reis RM, Ricardo S, Longatto-Filho A, Schmitt F, Baltazar F. Expression of monocarboxylate transporters 1, 2, and 4 in human tumours and their association with CD147 and CD44. J Biomed Biotechnol. (2010) 2010:427694. doi: 10.1155/2010/427694
27. Pinheiro C, Sousa B, Albergaria A, Paredes J, Dufloth R, Vieira D, et al. GLUT1 and CAIX expression profiles in breast cancer correlate with adverse prognostic factors and MCT1 overexpression. Histol Histopathol. (2011) 26:1279–86. doi: 10.14670/HH-26.1279
28. Pinheiro C, Longatto-Filho A, Scapulatempo C, Ferreira L, Martins S, Pellerin L, et al. Increased expression of monocarboxylate transporters 1, 2, and 4 in colorectal carcinomas. Virchows Arch. (2008) 452:139–46. doi: 10.1007/s00428-007-0558-5
29. Pinheiro C, Granja S, Longatto-Filho A, Faria AM, Fragoso M, Lovisolo SM, et al. GLUT1 expression in pediatric adrenocortical tumors: a promising candidate to predict clinical behavior. Oncotarget. (2017) 8:63835–45. doi: 10.18632/oncotarget.19135
30. Vander Heiden MG, DeBerardinis RJ. Understanding the intersections between metabolism and cancer biology. Cell. (2017) 168:657–69. doi: 10.1016/j.cell.2016.12.039
31. Ganapathy V, Thangaraju M, Prasad PD. Nutrient transporters in cancer: relevance to Warburg hypothesis and beyond. Pharmacol Ther. (2009) 121:29–40. doi: 10.1016/j.pharmthera.2008.09.005
32. Wykoff CC, Beasley NJ, Watson PH, Turner KJ, Pastorek J, Sibtain A, et al. Hypoxia-inducible expression of tumor-associated carbonic anhydrases. Cancer Res. (2000) 60:7075–83.
33. Smallbone K, Gavaghan DJ, Gatenby RA, Maini PK. The role of acidity in solid tumour growth and invasion. J Theor Biol. (2005) 235:476–84. doi: 10.1016/j.jtbi.2005.02.001
34. Chiche J, Ilc K, Brahimi-Horn MC, Pouyssegur J. Membrane-bound carbonic anhydrases are key pH regulators controlling tumor growth and cell migration. Adv Enzyme Regul. (2010) 50:20–33. doi: 10.1016/j.advenzreg.2009.10.005
35. Shin HJ, Rho SB, Jung DC, Han IO, Oh ES, Kim JY. Carbonic anhydrase IX (CA9) modulates tumor-associated cell migration and invasion. J Cell Sci. (2011) 124 (Pt 7):1077–87. doi: 10.1242/jcs.072207
36. Kalavska K, Chovanec M, Zatovicova M, Takacova M, Gronesova P, Svetlovska D, et al. Prognostic value of serum carbonic anhydrase IX in testicular germ cell tumor patients. Oncol Lett. (2016) 12:2590–8. doi: 10.3892/ol.2016.5010
37. Kalavska K, Cierna Z, Chovanec M, Takacova M, Svetlovska D, Miskovska V, et al. Prognostic value of intratumoral carbonic anhydrase IX expression in testicular germ cell tumors. Oncol Lett. (2017) 13:2177–85. doi: 10.3892/ol.2017.5745
38. Marini C, Salani B, Massollo M, Amaro A, Esposito AI, Orengo AM, et al. Direct inhibition of hexokinase activity by metformin at least partially impairs glucose metabolism and tumor growth in experimental breast cancer. Cell Cycle. (2013) 12:3490–9. doi: 10.4161/cc.26461
39. Patra KC, Wang Q, Bhaskar PT, Miller L, Wang Z, Wheaton W, et al. Hexokinase 2 is required for tumor initiation and maintenance and its systemic deletion is therapeutic in mouse models of cancer. Cancer Cell. (2013) 24:213–28. doi: 10.1016/j.ccr.2013.06.014
40. Sato-Tadano A, Suzuki T, Amari M, Takagi K, Miki Y, Tamaki K, et al. Hexokinase II in breast carcinoma: a potent prognostic factor associated with hypoxia-inducible factor-1alpha and Ki-67. Cancer Sci. (2013) 104:1380–8. doi: 10.1111/cas.12238
41. Tseng PL, Chen CW, Hu KH, Cheng HC, Lin YH, Tsai WH, et al. The decrease of glycolytic enzyme hexokinase 1 accelerates tumor malignancy via deregulating energy metabolism but sensitizes cancer cells to 2-deoxyglucose inhibition. Oncotarget. (2018) 9:18949–69. doi: 10.18632/oncotarget.24855
42. Katagiri M, Karasawa H, Takagi K, Nakayama S, Yabuuchi S, Fujishima F, et al. Hexokinase 2 in colorectal cancer: a potent prognostic factor associated with glycolysis, proliferation and migration. Histol Histopathol. (2017) 32:351–60. doi: 10.14670/HH-11-799
43. Wolf A, Agnihotri S, Micallef J, Mukherjee J, Sabha N, Cairns R, et al. Hexokinase 2 is a key mediator of aerobic glycolysis and promotes tumor growth in human glioblastoma multiforme. J Exp Med. (2011) 208:313–26. doi: 10.1084/jem.20101470
44. Kwee SA, Hernandez B, Chan O, Wong L. Choline kinase alpha and hexokinase-2 protein expression in hepatocellular carcinoma: association with survival. PLoS ONE. (2012) 7:e46591. doi: 10.1371/journal.pone.0046591
45. Guzman G, Chennuri R, Chan A, Rea B, Quintana A, Patel R, et al. Evidence for heightened hexokinase II immunoexpression in hepatocyte dysplasia and hepatocellular carcinoma. Dig Dis Sci. (2015) 60:420–6. doi: 10.1007/s10620-014-3364-3
46. Ogawa H, Nagano H, Konno M, Eguchi H, Koseki J, Kawamoto K, et al. The combination of the expression of hexokinase 2 and pyruvate kinase M2 is a prognostic marker in patients with pancreatic cancer. Mol Clin Oncol. (2015) 3:563–71. doi: 10.3892/mco.2015.490
47. Anderson M, Marayati R, Moffitt R, Yeh JJ. Hexokinase 2 promotes tumor growth and metastasis by regulating lactate production in pancreatic cancer. Oncotarget. (2017) 8:56081–94. doi: 10.18632/oncotarget.9760
48. Wang L, Xiong H, Wu F, Zhang Y, Wang J, Zhao L, et al. Hexokinase 2-mediated Warburg effect is required for PTEN- and p53-deficiency-driven prostate cancer growth. Cell Rep. (2014) 8:1461–74. doi: 10.1016/j.celrep.2014.07.053
49. Pedersen PL. Warburg, me and Hexokinase 2: Multiple discoveries of key molecular events underlying one of cancers' most common phenotypes, the “Warburg Effect”, i.e., elevated glycolysis in the presence of oxygen. J Bioenerg Biomembr. (2007) 39:211–22. doi: 10.1007/s10863-007-9094-x
50. Klepinin A, Ounpuu L, Mado K, Truu L, Chekulayev V, Puurand M, et al. The complexity of mitochondrial outer membrane permeability and VDAC regulation by associated proteins. J Bioenerg Biomembr. (2018) 50:339–54. doi: 10.1007/s10863-018-9765-9
51. Roberts DJ, Miyamoto S. Hexokinase II integrates energy metabolism and cellular protection: Akting on mitochondria and TORCing to autophagy. Cell Death Differ. (2015) 22:364. doi: 10.1038/cdd.2014.208
52. Uhlen M, Fagerberg L, Hallstrom BM, Lindskog C, Oksvold P, Mardinoglu A, et al. Proteomics. Tissue-based map of the human proteome. Science. (2015) 347:1260419. doi: 10.1126/science.1260419
53. Postic C, Leturque A, Printz RL, Maulard P, Loizeau M, Granner DK, et al. Development and regulation of glucose transporter and hexokinase expression in rat. Am J Physiol. (1994) 266:E548–59. doi: 10.1152/ajpendo.1994.266.4.E548
54. Sperger JM, Chen X, Draper JS, Antosiewicz JE, Chon CH, Jones SB, et al. Gene expression patterns in human embryonic stem cells and human pluripotent germ cell tumors. Proc Natl Acad Sci USA. (2003) 100:13350–5. doi: 10.1073/pnas.2235735100
55. Mathieu J, Zhang Z, Nelson A, Lamba DA, Reh TA, Ware C, et al. Hypoxia induces re-entry of committed cells into pluripotency. Stem Cells. (2013) 31:1737–48. doi: 10.1002/stem.1446
56. Dodo M, Kitamura H, Shima H, Saigusa D, Wati SM, Ota N, et al. Lactate dehydrogenase C is required for the protein expression of a sperm-specific isoform of lactate dehydrogenase a. J Biochem. (2018) 165:323–34. doi: 10.1093/jb/mvy108
57. Goldberg E. Lactate dehydrogenases and malate dehydrogenases in sperm: studied by polyacrylamide gel electrophoresis. Ann N Y Acad Sci. (1964) 121:560–70.
58. Zinkham WH, Blanco A, Clowry LJJr. An unusual isozyme of lactate dehydrogenase in mature testes: localization, ontogeny, and kinetic properties. Ann N Y Acad Sci. (1964) 121:571–88.
59. Tang H, Duan C, Bleher R, Goldberg E. Human lactate dehydrogenase A (LDHA) rescues mouse Ldhc-null sperm function. Biol Reprod. (2013) 88:96. doi: 10.1095/biolreprod.112.107011
60. Pelicano H, Martin DS, Xu RH, Huang P. Glycolysis inhibition for anticancer treatment. Oncogene. (2006) 25:4633–46. doi: 10.1038/sj.onc.1209597
61. Salani B, Marini C, Rio AD, Ravera S, Massollo M, Orengo AM, et al. Metformin impairs glucose consumption and survival in Calu-1 cells by direct inhibition of hexokinase-II. Sci Rep. (2013) 3:2070. doi: 10.1038/srep02070
62. Agnihotri S, Mansouri S, Burrell K, Li M, Mamatjan Y, Liu J, et al. Ketoconazole and posaconazole selectively target HK2-expressing glioblastoma cells. Clin Cancer Res. (2019) 25:844–55. doi: 10.1158/1078-0432.CCR-18-1854
Keywords: immunohistochemistry, metabolic reprogramming, testicular germ cell tumors, testicular neoplasms, Warburg effect
Citation: Bonatelli M, Silva ECA, Cárcano FM, Zaia MG, Lopes LF, Scapulatempo-Neto C and Pinheiro C (2019) The Warburg Effect Is Associated With Tumor Aggressiveness in Testicular Germ Cell Tumors. Front. Endocrinol. 10:417. doi: 10.3389/fendo.2019.00417
Received: 05 March 2019; Accepted: 10 June 2019;
Published: 28 June 2019.
Edited by:
Ugo De Giorgi, Romagnolo Scientific Institute for the Study and Treatment of Tumors (IRCCS), ItalyReviewed by:
Michal Mego, Campus Bio-Medico University, ItalyCopyright © 2019 Bonatelli, Silva, Cárcano, Zaia, Lopes, Scapulatempo-Neto and Pinheiro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Céline Pinheiro, Y2VsaW5lcGluaGVpcm9AZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.