
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cardiovasc. Med. , 19 March 2020
Sec. Atherosclerosis and Vascular Medicine
Volume 7 - 2020 | https://doi.org/10.3389/fcvm.2020.00031
This article is part of the Research Topic Established and Novel Roles of Platelets in Health and Disease View all 20 articles
 Idrissa Diallo1,2
Idrissa Diallo1,2 Abderrahim Benmoussa1,2Jonathan Laugier1,2
Abderrahim Benmoussa1,2Jonathan Laugier1,2 Abdimajid Osman3,4
Abdimajid Osman3,4 Walter E. Hitzler5
Walter E. Hitzler5 Patrick Provost1,2*
Patrick Provost1,2*Despite improvements in donor screening and increasing efforts to avoid contamination and the spread of pathogens in clinical platelet concentrates (PCs), the risks of transfusion-transmitted infections remain important. Relying on an ultraviolet photo activation system, pathogen reduction technologies (PRTs), such as Intercept and Mirasol, utilize amotosalen, and riboflavin (vitamin B2), respectively, to mediate inactivation of pathogen nucleic acids. Although they are expected to increase the safety and prolong the shelf life of clinical PCs, these PRTs might affect the quality and function of platelets, as recently reported. Upon activation, platelets release microparticles (MPs), which are involved in intercellular communications and regulation of gene expression, thereby mediating critical cellular functions. Here, we have used small RNA sequencing (RNA-Seq) to document the effect of PRT treatment on the microRNA profiles of platelets and derived MPs. PRT treatment did not affect the microRNA profile of platelets. However, we observed a specific loading of certain microRNAs into platelet MPs, which was impaired by treatment with Intercept or its Additive solution (SSP+). Whereas, Intercept had an impact on the microRNA profile of platelet-derived MPs, Mirasol did not impact the microRNA profile of platelets and derived MPs, compared to non-treated control. Considering that platelet MPs are able to transfer their microRNA content to recipient cells, and that this content may exert biological activities, those findings suggest that PRT treatment of clinical PCs may modify the bioactivity of the platelets and MPs to be transfused and argue for further investigations into PRT-induced changes in clinical PC content and function.
Derived from giant megakaryocytes in the hematopoietic bone marrow, platelets are small discoid and anucleate blood elements (1). Platelets have a key hemostatic role and are guardians of the structural integrity of blood vessels (2). Upon activation, platelets release extracellular vesicles (EVs), known as microparticles (MPs) (3), along with other particles, such as free mitochondria and mito-MPs (4). These MPs can mediate intercellular communications through delivery of bioactive molecules (5). With their rich content in protein-coding messenger RNAs, non-coding RNAs (e.g., microRNAs), cytokines, and lipids, platelet MPs may play an important role in gene regulation and homeostasis (5, 6). We reported previously that MPs released from human platelets can transfer functional microRNAs to human endothelial cells (1) and macrophages (2), in which platelet-derived microRNAs could regulate host cell gene expression.
Transfusion of clinical platelet concentrates (PCs) is required in various pathological conditions (e.g., thrombocytopenia, cancers, transplant surgery) (7) or for prophylactic purposes (8). In contrast to other blood products, clinical PCs, once collected and prepared (e.g., by apheresis or from buffy coats) are stored at room temperature under gentle agitation. This almost systematic procedure limits platelet storage to 5–7 days (9).
To increase their shelf life, these clinical PCs go through a series of routine tests and are sometimes treated with pathogen reduction technologies (PRTs) to eliminate the potential risks of viral, bacterial, fungal, or protozoal type contamination (10). The main PRTs available on the market are based on a photochemical principle (excitation at a given wavelength) to cross-link the pathogens' nucleic acids. The pathogen inhibitor Mirasol uses riboflavin (vitamin B2), a natural photochemical compound that binds to nucleic acids after UV exposure. This is followed by oxidation of guanine bases leading to single-strand breaks (11, 12). On the same principle, Intercept, along with its Additive solution, uses the properties of amotosalen to irreversibly block the replication of DNA and RNA, thus preventing the proliferation of pathogens (11, 13, 14). There are other pathogen inhibitors under development, such as Theraflex, which does not use photochemical compounds, but only ultraviolet C (UVC) irradiation (15).
However, these technologies, which are essentially based on photochemistry, do not receive unanimous agreement regarding their innocuousness on platelets and derived MPs, especially on their repertoire of functional nucleic acids (16). Although PRTs are convenient to reduce the risk of transfusion-transmitted infections, they can affect platelets and derived MPs' properties and, therefore, influence their functions. In fact, PRTs affect the metabolic (pH, sugars, nucleosides) and physiological parameters of platelets (e.g., increased platelet activation, reduced aggregation in response to agonists such as collagen, ADP, and thrombin, and increased risk of bleeding, as reported in several clinical cases) (17–20). However, less is known about their effects on the generation, content, and bioactivity of platelet MPs.
The advent of PRTs predated the discovery of the rich and functional nucleic acid content (e.g., messenger RNA, transfer RNA, microRNA, and long non-coding RNA) of platelets and derived MPs. Their effects on the profile of mRNAs and, in particular, microRNAs are yet to be unveiled. We have previously reported that PRT treatment of clinical PCs altered the level of some platelet microRNAs (21), suggesting that it might also alter the microRNA content of MPs derived from PRT-treated platelets. In this study, we have used small RNA sequencing (RNA-Seq) to characterize the microRNA profile of platelets, treated or not with pathogen inhibitors Mirasol or Intercept, as well as that of the MPs derived from these platelets.
The study was designed as previously described (19, 21). Briefly, clinical PCs were prepared from blood by apheresis using a standard blood bank protocol (19). PCs were subjected to one of the following four conditions: (1) Control (platelets stored in donor plasma); (2) Mirasol [platelets stored in donor plasma and treated with riboflavin and ultraviolet B (UVB) light]; (3) Additive solution [platelets stored in 65% storage solutions for platelets (SSP+; MacoPharma) and 35% donor plasma]; or (4) Intercept [platelets stored in SSP+ (Additive solution) and treated with amotosalen and UVA light]. For each group, 6 PC samples were pooled (n = 6 PCs per treatment, 24 samples in total, 4 pools). All PR treatments were performed according to the standard blood bank procedures or the manufacturer's instructions without modification.
Clinical PCs, treated or not with PRTs, or with the Additive solution, were stored under blood bank conditions (at room temperature under gentle rocking) for 7 days.
For platelet analysis, platelets were isolated from the PCs, as previously described (19), using anti-CD45 magnetic beads to minimize leukocyte co-isolation (<1 leukocyte per 3.2 million platelets; <0.03% of the platelet RNA preparations) (22, 23).
For MP analysis, platelets were sedimented at 1,000 g for 10 min. Platelet-free supernatant (PFS) was further spun a 18,000 g for 90 min to isolate platelet MPs.
Considering the role of MPs in intercellular communications, we analyzed the microRNA content of platelet MPs by RNA-Seq, as we did for platelets.
Total RNA from platelets or MPs was isolated using Trizol LS (Life Technologies—Ambion, Thermo-Fisher Scientific) and suspended in DEPC-treated DNase-RNase-free water (Invitrogen) prior to RNA purification and subsequent on-column DNase treatment using RNeasy mini-kit (Qiagen) following the manufacturer's protocol. Total RNA was then shipped on dry ice to the sequencing platform (ArrayStar).
The purity, quality, and concentration of total RNA samples were determined using NanoDrop ND-1000 (Thermo Scientific) and 2100 Bioanalyzer (Agilent). Total RNA of each sample was used to prepare the microRNA sequencing library, which included the following steps: (1) 3′-adapter ligation, (2) 5′-adapter ligation, (3) cDNA synthesis, (4) PCR amplification, and (5) size selection of PCR amplified fragments of ~130–150 base pairs (corresponding to small RNAs of ~15–35 nucleotides). The complete libraries were quantified by Agilent 2100 Bioanalyzer.
The samples were diluted to a final concentration of 8 pM and denatured as single-stranded DNA prior to cluster generation performed on an Illumina cBot using a TruSeq Rapid SR cluster kit (#GD-402-4001, Illumina). The clusters were then sequenced for 51 cycles on an Illumina HiSeq 2000 using TruSeq Rapid SBS Kits (#FC-402-4002, Illumina), as per the manufacturer's instructions.
The clean reads that passed the quality filter were processed to remove the adaptor sequence to generate the trimmed reads. All analyses displayed here were obtained from ArrayStar standard analysis pipeline and refined using R (Free Software Foundation). MicroRNA read counts were normalized as read counts per million microRNA alignments (RPM). Sequences known to be contaminant confounders from RNA isolation procedures were discarded before analysis.
Figures displayed in this manuscript were generated using R (Free Software Foundation), Inkscape software (Free Software Foundation) and/or Prism 8 (GraphPad Software, Inc.).
We first analyzed, by RNA-Seq, the profile of microRNAs in platelets either not treated (Control) or treated with Mirasol, Additive solution, or Intercept. MicroRNA profiling and clustering data suggest that treatment of platelets with Mirasol, Additive solution, or Intercept altered the global profile of platelet microRNAs (Figure 1A). However, when looking at the 20 most abundant platelet microRNAs, we obtained similar profiles between the four experimental conditions (Figure 1B).
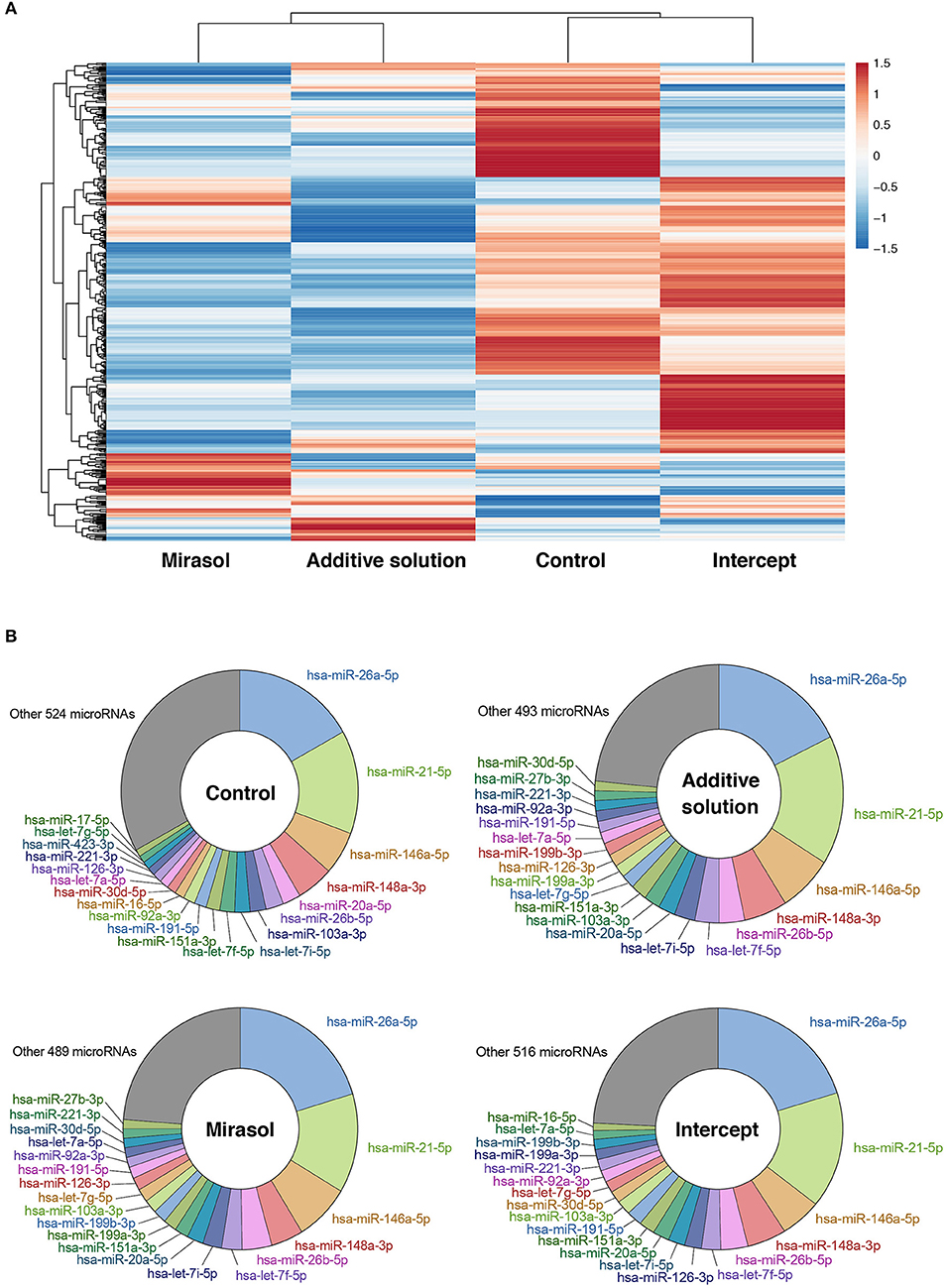
Figure 1. MicroRNA profile of PRT-treated human platelets. Clinical PCs were either left untreated in donor plasma (Control group) or treated with PRT agents Mirasol or Intercept, or with the Additive solution, used as the vehicle control for Intercept. Platelets were isolated from the clinical PCs, and total RNA was extracted and pooled from six donors per group prior to small RNA-Seq analysis. (A) Heatmap representation of the platelet microRNA profiles with clustering analysis, where fold changes vs. control PCs are indicated in graded color intensity. (B) Circular diagram representing the 20 most abundant microRNAs in each pooled sample, along with other microRNAs and their relative proportion. PCs, platelet concentrates; PRT, pathogen reduction technologies.
We observed that the 20 most abundant microRNAs, in both platelets (Figure 1B) and derived MPs (Figure 2A), comprised approximately two-thirds of all sequences. Fourteen of the 20 most abundant microRNAs in platelets were found among the 20 most abundant microRNAs in platelet MPs, suggesting that the microRNA profile of platelet MPs slightly differs from that of the platelets from which they derive (Figure 2A vs. Figure 1B).
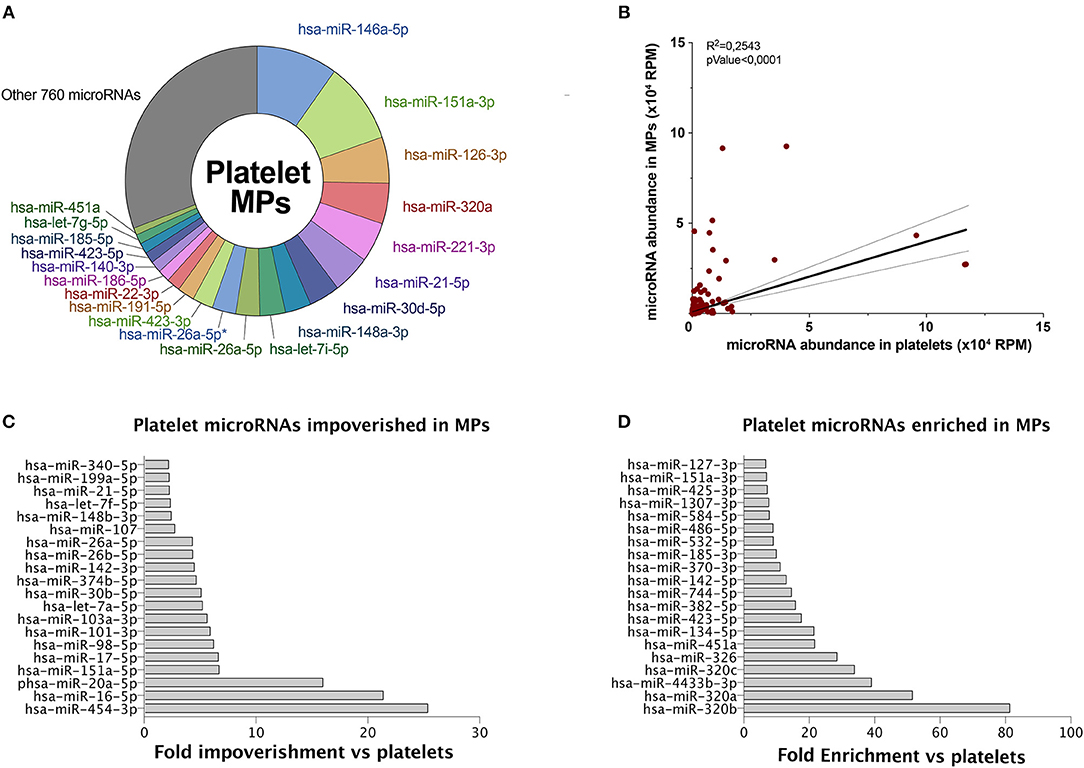
Figure 2. Platelet microRNAs are differentially partitioned into MPs. MPs were isolated from control PCs. Total RNA was extracted and pooled from six donors prior to small RNA-Seq analysis, as done for platelets in Figure 1. (A) Circular diagram representing the 20 most abundant microRNAs in platelet MPs, along with other microRNAs and their relative proportion. *IsomiR sequence. (B) Pearson's correlation of microRNA abundance in platelet MPs vs. platelets. (C) The 20 most abundant microRNAs, above 1,000 RPM, specifically impoverished in platelet MPs vs. platelets. (D) The 20 most abundant microRNAs, above 1,000 RPM, specifically enriched in platelet MPs vs. platelets. Hsa, Homo sapiens; MPs, microparticles; PCs, platelet concentrates; RPM, reads per million.
Platelet MPs (780 unique sequences; Figure 2A) appeared to contain a more diverse array of microRNAs than platelets (544 unique sequences; Figure 1B), with the difference concerning mainly the least abundant sequences.
A correlation of the platelet and derived MP microRNAs suggests that some microRNAs may be specifically loaded in platelet MPs (Figure 2B). Among the top 20 microRNAs above 1,000 reads per million (RPM), hsa-miR-454-3p, hsa-miR-16-5p, and hsa-miR-20a-5p are the three microRNAs most specifically enriched in platelets (versus MPs) (Figure 2C). For platelet MPs, the top three specific microRNAs were hsa-miR-320b, hsa-miR-320a, and hsa-miR-4433b-3p (Figure 2D).
Linear regression analyses unveiled that Mirasol treatment of platelets had no effect on the microRNA content of the released MPs (Figure 3A). Whereas, the exposure of platelets to the vehicle of Intercept, Additive solution, slightly disturbed the MP microRNA correlation with MPs released from control platelets (Figure 3B), the treatment of platelets with Intercept had more profound effects (Figure 3C). A closer examination of Figure 3C revealed two groups of microRNAs: those that are enriched (the steepest slope) and those that are impoverished (the smoothest slope) in MPs released from Intercept-treated platelets. These results suggest that Intercept may interfere with the natural process underlying MP formation and released from platelets, leading to the production of MPs with a different content and, possibly, different bioactivity.
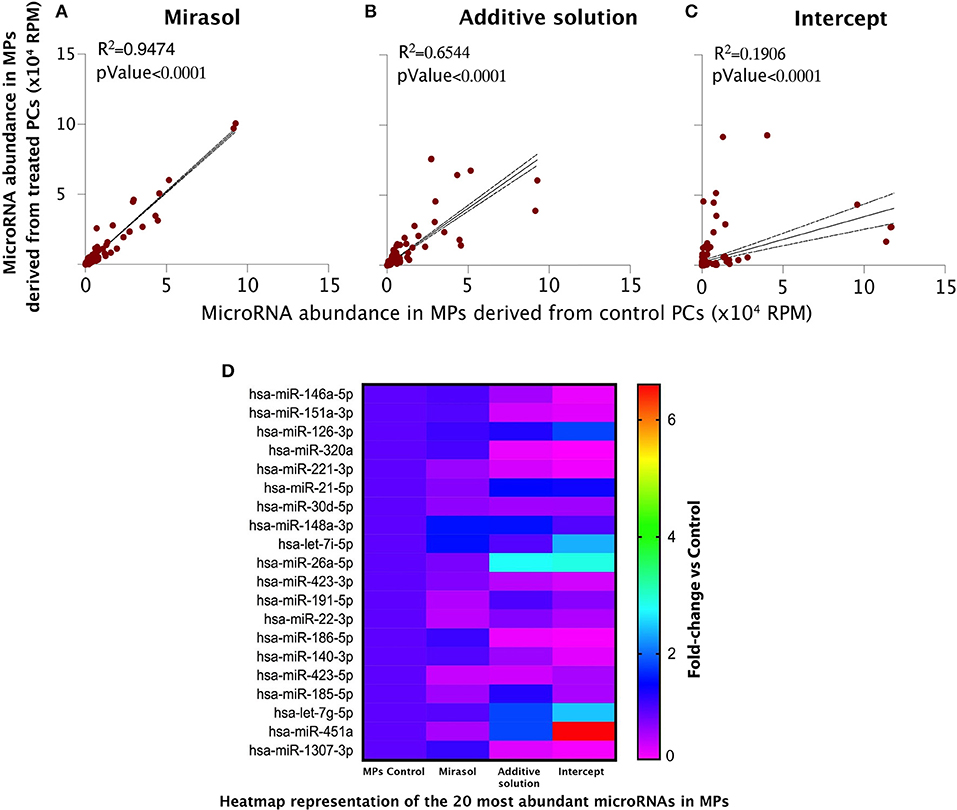
Figure 3. MicroRNA profile of the MPs derived from human platelets subjected to PRT treatment. MPs were isolated from PCs that were either left untreated in donor plasma (Control group) or treated with PRT agents Mirasol or Intercept, or with the Additive solution, used as the vehicle control for Intercept. (A–C) Pearson's correlation of the microRNA abundance in platelet MPs isolated from Mirasol-treated platelets (A), Additive solution-treated platelets (B), or Intercept-treated platelets (C) vs. control MPs. MPs, microparticles; RPM, reads per million. (D) Heatmap representation of the 20 most abundant microRNAs in MPs, where fold changes vs. control PCs are indicated in graded color intensity. MPs, microparticles; PCs, platelet concentrates; PRT, pathogen reduction technologies; RPM, reads per million.
We observed the same trend when focusing our analysis to the 20 most abundant microRNAs (Figure 3D and Tables 1–3). Mirasol treatment led to a feeble decrease in the level of certain microRNAs, such as hsa-miR-22-3p or hsa-miR-423-5p, in MPs compared to control MPs (Figure 3D and Table 1). On the contrary, the Additive solution decreased the level of certain highly abundant microRNAs, such as hsa-miR-151a-3p, hsa-miR-320a, or hsa-miR-221-3p, while increasing specifically the level of hsa-miR-26a-5p (Figure 3D and Table 2). Intercept treatment led to a more pronounced alteration of MP microRNA content, with a decrease in hsa-miR-185-5p levels and an increased abundance of hsa-let7i-5p (up to 3 fold) and hsa-miR-451a (up to 6.3 fold), compared to its vehicle (Additive solution) (Figure 3D and Table 3).

Table 1. Impact of Mirasol treatment on the microRNA profile of platelet-derived MPs.

Table 2. Impact of Additive solution treatment on the microRNA profile of platelet-derived MPs.
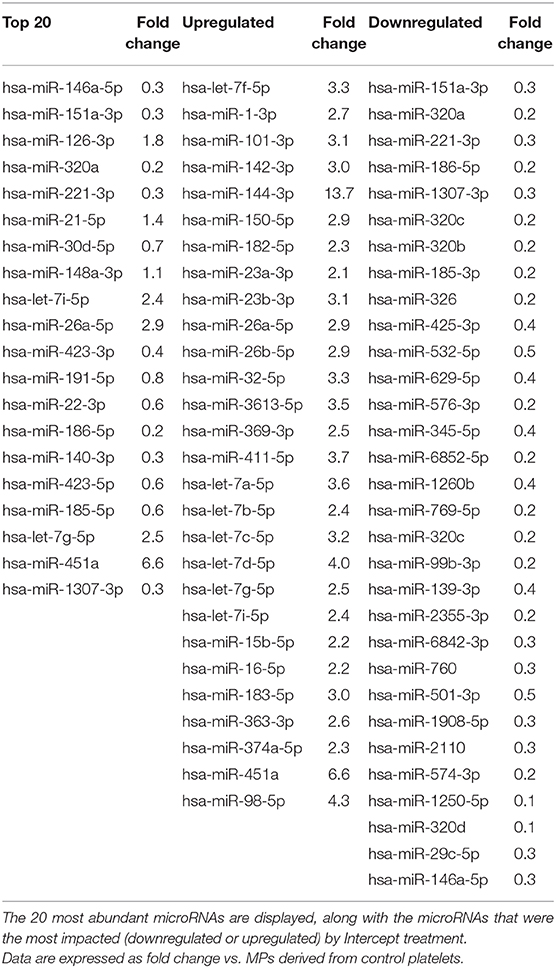
Table 3. Impact of Intercept treatment on the microRNA profile of platelet-derived MPs.
Similar trends were observed when analyzing the data of the other, less abundant microRNAs (not in the top 20, but >1,000 RPM). The effect of Mirasol was very limited, consisting mainly in downregulation of some microRNAs (Table 1). As above, the Additive solution modified (decreased or increased) the level of several other MP microRNAs of lower abundance (Table 2). Similarly, Intercept downregulated some MP microRNAs, such as hsa-miR-151a-3p (0.3-fold change vs. control MPs), while increasing the level of other MP microRNAs, such as hsa-miR-144-3p (13.7-fold change vs. control MPs) (Table 3).
Overall, our results indicate that Mirasol treatment of platelets had no marked effect on the microRNA profile of derived MPs, while Intercept exposure induced a more pronounced alteration in microRNA levels in platelet-derived MPs, an effect to which its Additive solution vehicle likely contributes.
Storage and quality control of PCs is a major challenge in blood banks, for which several checkpoints are set up from blood/platelet collection to PC transfusion. Based on the use of photochemical products designed to interfere with the genetic material of pathogens, through chemically induced cross-linking of nucleic acids, PRTs had revolutionized the control of PC safety and prolonged their shelf life. These technologies, however, were developed before we learned about the existence of functional nucleic acids (e.g., microRNAs) in human platelets (22, 24). Indeed, platelets carry all the components of the transcription (25) and translation (26) machinery of classical cells. They also contain a wide range of non-coding RNAs, including microRNAs (25, 27). Therefore, we cannot exclude the possibility that PRTs might impact the functional activity and/or level of nucleic acids contained in platelets or its derived MPs.
In this study, we expanded our previous qPCR analyses of selected microRNAs (21) by performing small RNA-Seq microRNA analyses of platelets, treated or not with PRT. These analyses corroborated our previous findings that Intercept may alter microRNA levels in exposed platelets (21), in addition to modulating the platelet mRNA transcriptome (19), which suggest that these consequences might be related.
PRTs induced variations on less abundant microRNAs. While such differences might have limited functional consequences individually, it is important to note that microRNAs generally act in clusters (28) and that changes, even subtle, in the level of several microRNAs might have an impact on platelet function or physiological response to agonists (21).
In the current study, we observed that platelets and their MPs had a different microRNA profile, suggesting a specific microRNA loading into MPs upon platelet activation. This difference, as also suggested by Diehl et al. (29), reflects an active and selective mechanism for packaging platelet microRNAs in MPs, which may confer specific regulatory functions to MPs.
Like outer membrane vesicles from bacteria (30) or other EV-like exosomes in eukaryotes (31), the profile of the genetic material, embedded in EVs, often depends on the physiological context and EV formation mechanisms at play in the cell of origin. It is, therefore, of importance to assess the effects that PRTs can have on the microRNA profile of platelet MPs.
Although we detected important differences in the microRNA repertoire of MPs derived from platelets exposed to Intercept or its Additive solution vehicle, none of the PRT treatments influenced the profile of microRNA isoforms (also known as isomiRs; Supplementary File 1). These observations suggest that PRT may influence microRNA loading into MPs rather than microRNA processing events.
Several studies have reported reduced platelet function upon Intercept treatment (19, 21, 32–34). Intercept has been shown to induce activation of p38, diminish the level of CD42 (also known as GpIb, which, in its complex, allows platelet adhesion and platelet clogging) (35), and deregulate expression of the pro-survival gene Bcl-xl and anti-apoptotic genes (21). In addition, to modulate the platelet mRNA transcriptome (19), Intercept was reported to alter platelet characteristics (mean volume), activation level, and aggregation response to physiological agonists (21). As the loading of genetic material (e.g., microRNAs) and the process of MP formation can be affected upon alteration of these characteristics and properties, the effect of Intercept on the microRNA profile of platelet MPs may thus be the consequence of all these interferences.
Intercept, in particular, seems to trigger the specific overrepresentation of hsa-miR-451a. This microRNAs is known to modulate cytokines, such as Macrophage Migration Inhibitory factor (MIF), which plays essential roles in the immune system and cell growth (36–38). As platelet MPs are internalized and transmit their microRNA content to macrophages (6), the transfer of this specific MP-enriched microRNA may reprogram the macrophage function of the PC recipient.
Other microRNAs, such as miR-26a, are also overrepresented in platelet MPs upon Intercept and Additive solution treatments. MiR-26a is involved in the regulation of numerous target genes (e.g., PTEN, SMAD1, AKT, and MAP3K2/MEKK2) and was found to play a role in normal tissue growth and development (39). The increase in platelet MP miR-26a levels may thus affect endothelial cell growth, as platelet MPs transmit their microRNA content to such cells (5).
The changes in the level of certain microRNAs in MPs derived from platelets exposed to Intercept or its Additive solution vehicle, may have long-term, cumulative effects upon multiple transfusions.
In this study, while there was no major effect on platelets, we found that PRT may impact the microRNA profile of platelet-derived MPs and induced overrepresentation of certain microRNAs. As platelet MPs may reprogram the function of macrophages by transferring their content in functional microRNAs, such as miR-126-3p (6) or miR-223 in endothelial cells (5), treatments that impact microRNA loading into MPs will also impact their bioactivity. The long-term consequences of transfusing modified platelet MPs to patients, and their detrimental/beneficial effects on health remain to be determined, especially in patients receiving PC on a regular basis.
The datasets generated for this study can be found in NCBI under accession number GSE138735.
The study was conducted by following the German pharmaceutical law for assessment of the quality of platelet products produced for routine use in hemotherapy and was approved by the Ethics Committee of the Medical Association of Rheinland-Pfalz (Ethik-Kommission bei der Landesärztekammer Rheinland-Pfalz–EK LÄK RLP). All platelet donors gave written informed consent to participate to the study.
WH and PP conceived and coordinated the study, and designed and planned the experiments. WH coordinated the sampling and treatment of the platelet concentrates. ID led the project, performed some of the experiments, and wrote the first draft of the manuscript. AB, AO, and JL performed some experiments and/or analysis. PP reviewed and finalized the manuscript. All authors have reviewed and approved the final manuscript before submission.
AB received Ph.D. studentship award no. 262093 from the Fonds de Recherche du Québec—Santé (FRQ-S). This work was supported by Grant Nos. 286,777 and 327,364 from the Canadian Blood Services (CBS)/Canadian Institutes of Health Research (CIHR) Partnership–Blood Utilization and Conservation Initiative, via Health Canada (to PP). The views expressed herein do not necessarily represent the view of the federal government.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcvm.2020.00031/full#supplementary-material
1. Thon JN, and Italiano JE. Platelets: production, morphology and ultrastructure. Handb Exp Pharmacol. (2012) 210:3–22. doi: 10.1007/978-3-642-29423-5_1
2. Ho-Tin-Noé B, Demers M, Wagner DD. How platelets safeguard vascular integrity. J Thromb Haemost. (2011) 9(Suppl.1):56–65. doi: 10.1111/j.1538-7836.2011.04317.x
3. Boilard E, Blanco P, Nigrovic PA. Platelets: active players in the pathogenesis of arthritis and SLE. Nat Rev Rheumatol. (2012) 8:534–42. doi: 10.1038/nrrheum.2012.118
4. Boudreau LH, Duchez A-C, Cloutier N, Soulet D, Martin N, Bollinger J, et al. Platelets release mitochondria serving as substrate for bactericidal group IIA-secreted phospholipase A2 to promote inflammation. Blood. (2014) 124:2173–83. doi: 10.1182/blood-2014-05-573543
5. Laffont B, Corduan A, Plé H, Duchez AC, Cloutier N, Boilard E, et al. (2013). Activated platelets can deliver mRNA regulatory Ago2∢microRNA complexes to endothelial cells via microparticles. Blood. 122:253–61. doi: 10.1182/blood-2013-03-492801
6. Laffont B, Corduan A, Rousseau M, Duchez AC, Lee CHC, Boilard E, et al. Platelet microparticles reprogram macrophage gene expression and function. Thromb Haemost. (2016) 115:311–23. doi: 10.1160/th15-05-0389
7. Wandt H, Schäfer-Eckart K, Greinacher A. Platelet transfusion in hematology, oncology and surgery. Dtsch Arztebl Int. (2014) 111:809–15. doi: 10.3238/arztebl.2014.0809
8. Crighton GL, Estcourt LJ, Wood EM, Trivella M, Doree C, Stanworth S. A therapeutic-only versus prophylactic platelet transfusion strategy for preventing bleeding in patients with haematological disorders after myelosuppressive chemotherapy or stem cell transplantation. Cochrane Database Syst Rev. (2015) 11:CD010981. doi: 10.1002/14651858.CD010981.pub2
9. Slichter SJ, Corson J, Jones MK, Christoffel T, Pellham E, Bailey SL, et al. Exploratory studies of extended storage of apheresis platelets in a platelet additive solution (PAS). Blood. (2014) 123:271–80. doi: 10.1182/blood-2013-05-501247
10. Mundt JM, Rouse L, Van den Bossche J, Goodrich RP. Chemical and biological mechanisms of pathogen reduction technologies. Photochem Photobiol. (2014) 90:957–64. doi: 10.1111/php.12311
11. Kwon SY, Kim IS, Bae JE, Kang JW, Cho YJ, Cho NS, et al. Pathogen inactivation efficacy of Mirasol PRT System and Intercept Blood System for non-leucoreduced platelet-rich plasma-derived platelets suspended in plasma. Vox Sang. (2014) 107:254–60. doi: 10.1111/vox.12158
12. Marschner S, Goodrich R. Pathogen reduction technology treatment of platelets, plasma and whole blood using riboflavin and UV light. Transfus Med Hemother. (2011) 38:8–18. doi: 10.1159/000324160
13. Infanti L, Stebler C, Job S, Ruesch M, Gratwohl A, Irsch J, et al. Pathogen-inactivation of platelet components with the INTERCEPT blood systemTM: a cohort study. Transfus Apher Sci. (2011) 45:175–81. doi: 10.1016/j.transci.2011.07.013
14. Irsch J, Lin L. Pathogen inactivation of platelet and plasma blood components for transfusion using the INTERCEPT blood systemTM. Transfus Med Hemother. (2011) 38:19–31. doi: 10.1159/000323937
15. Sandgren P, Tolksdorf F, Struff WG, Gulliksson H. In vitro effects on platelets irradiated with short-wave ultraviolet light without any additional photoactive reagent using the THERAFLEX UV-Platelets method. Vox Sang. (2011) 101:35–43. doi: 10.1111/j.1423-0410.2010.01454.x
16. Magron A, Laugier J, Provost P, Boilard E. Pathogen reduction technologies: the pros and cons for platelet transfusion. Platelets. (2018) 29:2–8. doi: 10.1080/09537104.2017.1306046
17. Janetzko K, Cazenave J-P, Klüter H, Kientz D, Michel M, Beris P, et al. Therapeutic efficacy and safety of photochemically treated apheresis platelets processed with an optimized integrated set. Transfusion. (2005) 45:1443–52. doi: 10.1111/j.1537-2995.2005.00550.x
18. Kerkhoffs J-LH, van Putten WLJ, Novotny VMJ, Te Boekhorst PAW, Schipperus MR, Zwaginga JJ, et al. Clinical effectiveness of leucoreduced, pooled donor platelet concentrates, stored in plasma or additive solution with and without pathogen reduction. Br J Haematol. (2010) 150:209–17. doi: 10.1111/j.1365-2141.2010.08227.x
19. Osman A, Hitzler WE, Ameur A, Provost P. Differential expression analysis by RNA-seq reveals perturbations in the platelet mRNA transcriptome triggered by pathogen reduction systems. PLoS ONE. (2015) 10:e0133070. doi: 10.1371/journal.pone.0133070
20. Stivala S, Gobbato S, Infanti L, Reiner MF, Bonetti N, Meyer SC, et al. Amotosalen/ultraviolet A pathogen inactivation technology reduces platelet activatability, induces apoptosis and accelerates clearance. Haematologica. (2017) 102:1650–60. doi: 10.3324/haematol.2017.164137
21. Osman A, Hitzler WE, Meyer CU, Landry P, Corduan A, Laffont B, et al. Effects of pathogen reduction systems on platelet microRNAs, mRNAs, activation, and function. Platelets. (2015) 26:154–63. doi: 10.3109/09537104.2014.898178
22. Landry P, Plante I, Ouellet DL, Perron MP, Rousseau G, Provost P. Existence of a microRNA pathway in anucleate platelets. Nat Struct Mol Biol. (2009) 16:961–6. doi: 10.1038/nsmb.1651
23. Teruel-Montoya R, Kong X, Abraham S, Ma L, Kunapuli SP, Holinstat M, et al. MicroRNA expression differences in human hematopoietic cell lineages enable regulated transgene expression. PLoS ONE. (2014) 9:e102259. doi: 10.1371/journal.pone.0102259
24. Zimmerman GA, Weyrich AS. Signal-dependent protein synthesis by activated platelets: new pathways to altered phenotype and function. Arterioscler Thromb Vasc Biol. (2008) 28:s17–24. doi: 10.1161/ATVBAHA.107.160218
25. Schubert S, Weyrich AS, Rowley JW. A tour through the transcriptional landscape of platelets. Blood. (2014) 124:493–502. doi: 10.1182/blood-2014-04-512756
26. Weyrich AS, Dixon DA, Pabla R, Elstad MR, McIntyre TM, Prescott SM, et al. Signal-dependent translation of a regulatory protein, Bcl-3, in activated human platelets. Proc Natl Acad Sci USA. (1998) 95:5556–61. doi: 10.1073/pnas.95.10.5556
27. Plé H, Landry P, Benham A, Coarfa C, Gunaratne PH, Provost P. The repertoire and features of human platelet microRNAs. PLoS ONE. (2012) 7:e50746. doi: 10.1371/journal.pone.0050746
28. Yu J, Wang F, Yang GH, Wang FL, Ma YN, Du ZW, et al. Human microRNA clusters: genomic organization and expression profile in leukemia cell lines. Biochem Biophys Res Commun. (2006) 349:59–68. doi: 10.1016/j.bbrc.2006.07.207
29. Diehl P, Fricke A, Sander L, Stamm J, Bassler N, Htun N, et al. Microparticles: major transport vehicles for distinct microRNAs in circulation. Cardiovasc Res. (2012) 93:633–44. doi: 10.1093/cvr/cvs007
30. Ellis TN, Kuehn MJ. Virulence and immunomodulatory roles of bacterial outer membrane vesicles. Microbiol Mol Biol Rev. (2010) 74:81–94. doi: 10.1128/MMBR.00031-09
31. Hessvik NP, Llorente A. Current knowledge on exosome biogenesis and release. Cell Mol Life Sci. (2018) 75:193–208. doi: 10.1007/s00018-017-2595-9
32. Abonnenc M, Crettaz D, Tacchini P, Di Vincenzo L, Sonego G, Prudent M, et al. Antioxidant power as a quality control marker for completeness of amotosalen and ultraviolet A photochemical treatments in platelet concentrates and plasma units. Transfusion. (2016) 56:1819–27. doi: 10.1111/trf.13638
33. Johnson L, Hyland R, Tan S, Tolksdorf F, Sumian C, Seltsam A, et al. In vitro quality of platelets with low plasma carryover treated with ultraviolet C light for pathogen inactivation. Transfus Med Hemother. (2016) 43:190–7. doi: 10.1159/000441830
34. Picker SM, Oustianskaia L, Schneider V, Gathof BS. Functional characteristics of apheresis-derived platelets treated with ultraviolet light combined with either amotosalen-HCl (S-59) or riboflavin (vitamin B2) for pathogen-reduction. Vox Sang. (2009) 97:26–33. doi: 10.1111/j.1423-0410.2009.01176.x
35. Hechler B, Ravanat C, Gachet C. Amotosalen/UVA pathogen inactivation technology reduces platelet activability, induces apoptosis and accelerates clearance. Haematologica. (2017) 102:e502–3. doi: 10.3324/haematol.2017.180539
36. Graham A, Falcone T, Nothnick WB. The expression of microRNA-451 in human endometriotic lesions is inversely related to that of macrophage migration inhibitory factor (MIF) and regulates MIF expression and modulation of epithelial cell survival. Hum Reprod. (2015) 30:642–52. doi: 10.1093/humrep/dev005
37. Lolis E, Bucala R. Macrophage migration inhibitory factor. Expert Opin Ther Targets. (2003) 7:153–64. doi: 10.1517/14728222.7.2.153
38. Nishihira J. Macrophage migration inhibitory factor (MIF): its essential role in the immune system and cell growth. J Interferon Cytokine Res. (2000) 20:751–62. doi: 10.1089/10799900050151012
Keywords: clinical platelet concentrate, pathogen reduction, mirasol, intercept, extracellular vesicles, small RNA-sequencing, microRNA
Citation: Diallo I, Benmoussa A, Laugier J, Osman A, Hitzler WE and Provost P (2020) Platelet Pathogen Reduction Technologies Alter the MicroRNA Profile of Platelet-Derived Microparticles. Front. Cardiovasc. Med. 7:31. doi: 10.3389/fcvm.2020.00031
Received: 04 October 2019; Accepted: 21 February 2020;
Published: 19 March 2020.
Edited by:
Paul Jurasz, University of Alberta, CanadaReviewed by:
Vera Ignjatovic, Royal Children's Hospital, AustraliaCopyright © 2020 Diallo, Benmoussa, Laugier, Osman, Hitzler and Provost. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patrick Provost, cGF0cmljay5wcm92b3N0QGNyY2h1ZGVxdWViZWMudWxhdmFsLmNh
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.