 Rodnei Dennis Rossoni1,2
Rodnei Dennis Rossoni1,2 Patrícia Pimentel de Barros1,2
Patrícia Pimentel de Barros1,2 Iatã do Carmo Mendonça3
Iatã do Carmo Mendonça3 Rebeca Previate Medina3
Rebeca Previate Medina3 Dulce Helena Siqueira Silva3Beth Burgwyn Fuchs2
Dulce Helena Siqueira Silva3Beth Burgwyn Fuchs2 Juliana Campos Junqueira1*
Juliana Campos Junqueira1* Eleftherios Mylonakis2
Eleftherios Mylonakis2- 1Department of Biosciences and Oral Diagnosis, Institute of Science and Technology, São Paulo State University/UNESP, São José dos Campos, Brazil
- 2Division of Infectious Diseases, Rhode Island Hospital, Warren Alpert Medical School at Brown University, Providence, RI, United States
- 3Department of Organic Chemistry, Center for Bioassays, Biosynthesis and Ecophysiology of Natural Products, Institute of Chemistry, São Paulo State University, UNESP, Araraquara, Brazil
Candida auris has emerged as a medically important pathogen with considerable resistance to antifungal agents. The ability to produce biofilms is an important pathogenicity feature of this species that aids escape of host immune responses and antimicrobial agents. The objective of this study was to verify antifungal action using in vitro and in vivo models of the Lactobacillus paracasei 28.4 probiotic cells and postbiotic activity of crude extract (LPCE) and fraction 1 (LPF1), derived from L. paracasei 28.4 supernatant. Both live cells and cells free supernatant of L. paracasei 28.4 inhibited C. auris suggesting probiotic and postbiotic effects. The minimum inhibitory concentration (MIC) for LPCE was 15 mg/mL and ranges from 3.75 to 7.5 mg/mL for LPF1. Killing kinetics determined that after 24 h treatment with LPCE or LPF1 there was a complete reduction of viable C. auris cells compared to fluconazole, which decreased the initial inoculum by 1-logCFU during the same time period. LPCE and LPF1 significantly reduced the biomass (p = 0.0001) and the metabolic activity (p = 0.0001) of C. auris biofilm. There was also a total reduction (~108 CFU/mL) in viability of persister C. auris cells after treatment with postbiotic elements (p < 0.0001). In an in vivo study, injection of LPCE and LPF1 into G. mellonella larvae infected with C. auris prolonged survival of these insects compared to a control group (p < 0.05) and elicited immune responses by increasing the number of circulating hemocytes and gene expression of antimicrobial peptide galiomicin. We concluded that the L. paracasei 28.4 cells and postbiotic elements (LPCE and LPF1) have antifungal activity against planktonic cells, biofilms, and persister cells of C. auris. Postbiotic supplementation derived from L. paracasei 28.4 protected G. mellonella infected with C. auris and enhanced its immune status indicating a dual function in modulating a host immune response.
Introduction
Opportunistic infections are caused by non-pathogenic microorganisms which become pathogenic when the body's defense system is impaired (Riccardi et al., 2019). Candida spp. can cause vaginitis, oral candidiasis, cutaneous candidiasis, and candidemia. This genus are responsible the main opportunistic yeast infection in the world (Jacobsen et al., 2012; Wachtler et al., 2012; Martins et al., 2014).
Most of these infections originate from biofilms (Vicariotto et al., 2012; Matsubara et al., 2016a,b), defined as complex and dynamic structures consisting of cells encased in a matrix of extracellular polymeric substances (Serra et al., 2015; Rodrigues M. E. et al., 2019). The adhered cells of the biofilm are protected from the external environment by the extracellular matrix that shields them and contributes to the increased antimicrobial resistance. The development of Candida spp. biofilms is one of the main reasons that fungal cells exhibit resistance to antifungals (Davies, 2003; Rodrigues et al., 2016; Cernakova et al., 2019; Rodrigues M. E. et al., 2019). Thus, treatment of infections derived from biofilms represents an important challenge today.
C. auris is a pathogenic yeast associated with invasive infections and it has been reported from 32 countries across six continents within a decade (Chakrabarti and Singh, 2020). Previous studies suggest that this species is highly tolerant to thermal and osmotic stresses, possessing the ability to persistently colonize hospital environments, and thereby causing outbreaks (Rossato and Colombo, 2018; Spivak and Hanson, 2018; Sabino et al., 2020). Recently, the Centers for Disease Control and Prevention (CDC) published a notice to different public health officials informing them of C. auris isolation in U.S. patients (Centers for Disease Control and Prevention, 2016; Sekyere and Asante, 2018). The emergence of C. auris is concerning because this yeast has or may develop resistance to different antifungal agents (Lockhart et al., 2017; Chaabane et al., 2019), making infections difficult to treat. Lockhart et al. (2017) evaluated the antifungal susceptibility of 41 isolates of C. auris from 54 patients during 2012–2015. The authors found that 93 and 35% of C. auris isolates were resistant to fluconazole (FLC) and amphotericin B, respectively. In addition, difficulties in its identification in the routine diagnostic laboratory have a significant impact on outbreak detection and appropriate management (Bidaud et al., 2018).
With poor response to standard antimicrobials and the deficiency in developing new antifungal agents into the clinic, it stands that alternate means of treating the fungal diseases are warranted. In this context, the use of probiotics has been considered a promising strategy for prevention and control of several fungal infections (Mailander-Sanchez et al., 2012; Matsubara et al., 2016a; Hu et al., 2017; Rodrigues C. F. et al., 2019). Probiotics are live microorganisms or microbial cell components that beneficially affect host health (Guarner et al., 2012; Janczarek et al., 2016; Matsubara et al., 2016b; Rossoni et al., 2018), but their clinical application in immunocompromised patients is limited due to the possibility of bacteremia caused by bacteria of the genus Lactobacillus (Cannon et al., 2005; Salminen et al., 2006).
To address the risk for live cell probiotics potential of bacteremia, postbiotics have emerged based on the concept that bacterial viability is not essential for probiotics to exert beneficial effects on human health (Tsilingiri and Rescigno, 2013; Cicenia et al., 2014). Postbiotics are products of probiotic bacteria that have biological activity in the host (Mosca et al., 2019) and include metabolites, fractions of microbial cells, fatty acids, proteins, polysaccharides, cell lysates, peptidoglycan derived peptides, and structures responsible for adhesion, such as pili (Wegh et al., 2019). In previous studies by our research group, we isolated the Lactobacillus paracasei strain 28.4 from the oral cavity and showed its inhibitory activity against the commensal fungus C. albicans in both in vitro and in vivo models (Rossoni et al., 2017, 2018; de Barros et al., 2018; Santos et al., 2019).
In the present study, the supernatant of L. paracasei 28.4 was extracted and fractionated to determine if postbiotic elements could be effective at inhibiting the emergent species C. auris. Particularly, examining their effects on biofilm state that can plague immunocompromised patients that spur recurrent and invasive infections, also, we examined cell-free postbiotic supernatant inhibitory activity using the invertebrate infection model Galleria mellonella to look for host responses.
Materials and Methods
Strains
A panel of C. auris strains (designated AR-BANK#0381 to AR-BANK#0390) obtained from the antimicrobial resistance bank of the CDC was used in this study. The strains in this panel are listed in Table 1. L. paracasei 28.4, an isolate from the oral cavity of a caries-free individual, was used to obtain crude extract and supernatant fractions. Our research group isolated, identified, and characterized this probiotic strain demonstrating its antifungal action both in vitro and in vivo studies (Rossoni et al., 2017, 2018; de Barros et al., 2018; Santos et al., 2019; Ribeiro et al., 2020).

Table 1. C. auris isolates used in this study.
All C. auris isolates were cultured in 1% yeast extract, 2% peptone, 2% dextrose (YPD broth, Difco, Detroit, USA) for 24 h at 37°C, and L. paracasei 28.4 was grown on Man Rogosa and Sharpe (MRS) agar (Difco, Franklin Lakes, NJ, USA) for 48 h at 37°C (5% CO2).
In vitro Coinfection Cultures in a Planktonic Environment
Coinfection cultures were performed in 2 mL of BHI broth at 37°C in a rollerdrum according Peleg et al. (2008) with modifications. Standardized inoculants of C. auris (1 × 105 cells/mL) and L. paracasei 28.4 (1 × 108 cells/mL) were incubated together for 24, 48, and 72 h. At the indicated time points, the CFU were enumerated for each group. YPD plates containing kanamycin (45 μg/mL) and MRS plates containing fluconazole (32 μg/mL) were used to determine C. auris and L. paracasei CFUs, respectively. Results were obtained from three independent experiments.
L. paracasei 28.4 Postbiotic Elements: Supernatant Preparation, Extraction, and Fractionation
First, the L. paracasei 28.4 supernatant was produced according to Ribeiro et al. (2017). An inoculum of 1 mL of the standard suspension (107 cells/mL) was seeded into 6 mL of MRS broth and incubated at 37°C for 24 h under microaerophilic conditions. After this incubation, the broth was centrifuged (5,000 RPM for 10 min) and filtered with a 0.22 μm pore size membrane (MFS, Dublin, CA, USA).
Next, postbiotic elements were obtained according to Medina et al. (2019). In total, 4 L of supernatant were extracted with EtOAc (3 × 50% of each medium volume) for the extraction of the active compounds. After evaporation of EtOAc using a rotary evaporator (Buchi rotavapor, Buchi, São Paulo, Brazil), L. paracasei 28.4 crude extract (LPCE) was obtained (1.18 g). LPCE crude extract was analyzed using HPLC with two LC6AD pumps, CBM-20A communicator, SIL-10AF automatic injector, and SPD-M20A diode array detector (Shimadzu, Columbia, MD, USA), and the column used was a Luna Phenomenex octadecyl silane (C-18) analytical 250 × 4.6 mm, and gradient elution H2O /MeOH (95:05 → 0:100) for 45 min. LPCE was fractionated using 20 g of C18 silica in an open column in which methanol and water were employed as a stationary phase in a polarity gradient (26:74, 51:49, 75:25, and 90:10). Fraction 1 of the supernatant (LPF1) was used in all subsequent assays as well as the LPCE. An aliquot of LPCE and LPF1 were cultured in brain heart infusion (BHI) broth (Sigma, St. Louis, MO, USA) to ensure that there was no microbial growth in the postbiotic elements.
Minimum Inhibitory Concentration
To determine the minimal inhibitory concentration (MIC) of LPCE and LPF1 against the C. auris strains, colonies of each strain were inoculated in 5 mL of yeast peptone dextrose (YPD) media (Sigma, St. Louis, MO, USA) and grown overnight at 37°C. The cells were harvested by centrifugation at 5,000 rpm for 5 min and washed with phosphate-buffered saline (PBS). Subsequently, the cell pellets were suspended in RPMI 1640 medium (Sigma, St. Louis, MO, USA). The cell count was determined using a hemocytometer and adjusted to 1.0 × 103 cells/mL. Susceptibility patterns of the isolates to LPCE and LPF1 were determined by performing the broth microdilution assay. The final concentrations of LPCE and LPF1 ranged from 30 to 0.029 mg/mL. Fluconazole and amphotericin B (Sigma-Aldrich, St. Louis, MO, USA) were used as a positive control and the assay was performed according the Clinical and Laboratory Standards Institute (CLSI) document M27-A2 (National Committee for Clinical Laboratory Standards, 2002). The final concentrations of fluconazole and amphotericin B ranged from 64 to 0.125 μg/mL. The resistance breakpoints were used as described in the Clinical and Laboratory Standards Institute (CLSI) guidelines based on C. albicans interpretive breakpoints (Fluconazole: ≤ 8.0 μg /mL for susceptible, ≥64 μg /mL for resistant; Amphotericin B: >1 μg/ml for resistant) (National Committee for Clinical Laboratory Standards, 2002).
Minimum Fungicidal Concentration
The minimum fungicidal concentration (MFC) was determined as follows. In total, 10 μL of yeast culture from each microwell of the MIC assay was plated on YPD agar and incubated at 35°C overnight. The static/cidal parameter can be roughly estimated by comparing the MFC of a given antifungal to its MIC. If the MFC is ≤ 4 × MIC the drug is considered cidal (Pfaller et al., 2004).
Time to Kill Assays
After the MIC test of all C. auris strains, the CAU-01 strain was selected for the subsequent tests based on its sensitivity to fluconazole and amphotericin B. This strain showed the lowest MIC value for both antifungals, a requirement for inducing persister cells. Therefore, C. auris strain CAU-01 was explored to interrogate the killing effects of LPCE and LPF1. The assays were carried out in 10-mL tubes (BD Biosciences, San Diego, CA, USA) in triplicate according to Tharmalingam et al. (2019) with modifications. Briefly, log-phase cultures of CAU-01 were diluted in fresh RPMI medium to a density of 106 cells/mL LPCE and LPF1 were added at concentrations 3.75–120 mg/mL (corresponding to 1 × MIC−8 × MIC), and the tubes were incubated at 37°C with agitation (200 rpm). Portions of cell suspensions were withdrawn at predetermined time points (24, 48, and 72 h). Serial dilutions were plated on YPD agar to determine the colony-forming unit/mL (CFU/mL) of the cell suspensions. CFU were determined after incubation for 48 h at 37°C. Three independent experiments were performed. As a positive control, we included the antifungal agent fluconazole at 4, 16 and 32 μg/mL (corresponding to 1 × MIC−8 × MIC).
Biofilm Formation
Evaluation of biofilm formation was performed in 96-well microtiter plates (Corning, New York, NY, USA) following the methodology described by Vilela et al. (2015) and Rossoni et al. (2018), with some modifications. Briefly, 100 μL of C. auris standard suspensions (1.0 × 107 cells) were deposited into 96-well microtiter plates, after which the plates were placed on a 75-rpm shaking incubator at 37°C for 90 min. Each well was washed twice with PBS, and 200 μL of yeast nitrogen base (YNB) broth (Sigma, St. Louis, MO, USA) supplemented with 100 mM glucose with LPCE or LPF1 were added to the wells of each plate at the concentrations of 0.5 × MIC, 1 × MIC, and 2 × MIC. For the control groups, 200 μL of YNB broth supplemented with 100 mM glucose without LPCE or LPF1 was added. The plate was incubated for 48 h at 37°C with shaking at 75 rpm. The liquid medium was replaced after 24 h and the treated groups received fresh LPCE, LPF1, or fluconazole dilutions. After the incubation period, each well was washed twice with PBS for subsequent analysis of total biomass and metabolic activity. As a positive control, we included the antifungal agent fluconazole at 4 and 8 μg/mL (corresponding to 1 × MIC and 2 × MIC).
Analysis of Biofilms by Total Biomass Quantification
After biofilm formation, the biofilm biomass was quantified utilizing an assay previously described by Peeters et al. (2008), with modifications. For biofilm fixation, 100 μL of 99% methanol was added to the wells (Sigma-Aldrich, St. Louis, MO, USA). After 15 min, the supernatants were removed and the plates were air-dried. Then, 100 μL of a 1% crystal violet (CV) solution was added to all wells. After 20 min, the residual CV solution was removed by washing with PBS. Finally, bound CV was released by adding 150 μL of 33% acetic acid (Sigma-Aldrich, St. Louis, MO, USA). The absorbance was measured at 540 nm. All steps were carried out at room temperature. The CV assay was performed as two independent experiments with n = 6 biofilms per group.
Analysis of Biofilms by XTT Reduction Assay Colorimetric Assay
The biofilms formed also were evaluated by a metabolic assay based on the reduction of XTT, a tetrazolium salt (Sigma-Aldrich, St. Louis, MO, USA) according Jin et al. (2004) and Rossoni et al. (2019). Briefly, XTT salt was dissolved in PBS at a final concentration of 1 mg/mL. Immediately before each assay, a menadione (Sigma-Aldrich, St. Louis, MO, USA) solution was prepared at a final concentration of 0.4 mM and filter-sterilized. The XTT solution was thawed prior to each assay and mixed with the menadione solution at a ratio of 20:1 (v/v). Each well was washed two times with 200 μL of PBS to remove any non-adherent cells. Next, 158 μL of PBS, 40 μL of XTT, and 2 μL of menadione were added to each of the pre-washed wells. The plates were incubated in the dark at 37°C for 3 h. Afterwards, 100 μL of the solution was transferred to a new well, and any colorimetric change in the solution was measured using a microtiter plate reader (Tp Reader; Thermo Plate, Shenzhen, China) at 490 nm. The XTT assay was performed as two independent experiments with n = 6 biofilms per group.
Isolation and Susceptibility of C. auris Persisters Cells
For this study, the methodologies described by LaFleur et al. (2006) and Al-Dhaheri and Douglas (2008) were used with some modifications. Briefly, C. auris was grown for 72 h at 37°C with shaking in RPMI 1640 with L-glutamine and 0.165 M MOPS growth medium (50 mL in 250-ml flasks) to isolate persister cells in a stationary-phase cultures. After 72 h, cells from the stationary-phase cultures were harvested and washed twice in PBS. All cell suspensions were adjusted to concentrations of ~5 × 108 cells/mL. An aliquot of 1 mL of this suspension, containing 10x, 50x, 100x, 150x, and 200x MICs of indicated antifungals (amphotericin B and fluconazole), was added to the wells of a 2 mL deep well assay block (Corning Costar 3960) and incubated at 37°C, with shaking at 225 rpm for 48 h. In groups containing postbiotic elements, the stationary-phase culture suspension was treated with 10 × MIC of LPCE or LPF1. Control cells were treated similarly with buffered medium without antifungal agent. At designated times, 50 μL samples were removed, serially diluted, and spot-plated on YPD agar plates to enumerate the number of persister cells. This experiment was conducted in triplicate.
G. mellonella Survival
G. mellonella survival was evaluated following the methodologies described by Mylonakis et al. (2005) and Rossoni et al. (2017), with some modifications. G. mellonella (Vanderhorst Wholesale, St. Marys, OH, USA) in their final larval stage were stored in the dark and used within 7 days from shipment. Sixteen randomly chosen G. mellonella larvae with similar weight and size (250–350 mg) were used per group in all assays. Two control groups were included in the assays—one group was inoculated with phosphate-buffered saline (PBS), while the other group received no injection as a control to evaluate general viability.
In view of the lack of studies in the literature that used postbiotics elements on G. mellonella as an experimental model, the toxicity of LPCE and LPF1 was evaluated prior to the study of experimental candidiasis. The postbiotic elements were injected directly into the hemolymph of G. mellonella at varying concentrations (10–80 mg/kg), and survival curves were constructed. Larvae were incubated at 37°C and monitored daily for survival.
For the experimental candidiasis, C. auris suspension was prepared from cultures in 5 mL of YPD liquid medium and incubated at 37°C for 18 h. Afterwards, cells were centrifuged at 2,000 × g for 10 min, and the supernatant was discarded. The cell pellet was washed twice and suspended in PBS. Cell densities were adjusted using a hemocytometer.
To evaluate the effects of LPCE or LPF1 on C. auris infection, the larvae were pre-treated by injecting the LPCE (80 mg/kg) or LPF1 (30 mg/kg) through the last left proleg (volume, 10 μL). After 2 h, larvae were infected with 106 cells/larvae of C. auris suspended in PBS at the last right proleg (10 μL of volume). Larvae were incubated at 37°C and monitored daily for survival. Larvae were considered dead when they displayed no movement in response to touch.
Quantification of G. mellonella Hemocytes
In order to investigate the immunological mechanisms associated with the postbiotics elements against infection by C. auris, larvae were pre-treated with LPCE (80 mg/kg) or LPF1 (30 mg/kg) and infected with C. auris as described above. Hemocytes were collected from the hemocoel at 4 h post-injection with C. auris. Larvae were bled into tubes containing cold, sterile insect physiologic saline (IPS) (150 mM sodium chloride; 5 mM potassium chloride; 100 mM Tris—hydrochloride, pH 6.9 with 10 mM EDTA, and 30 mM sodium citrate). The hemocytes were identified based on cell morphology and quantified using a hemocytometer. The results were averaged from four replicates.
Analysis of Peptide Expression
G. mellonella gene expression was evaluated following the methodologies described by Mowlds et al. (2010) and Rossoni et al. (2017). After pre-treatments and infection, larval RNA was extracted using TRIzol (Ambion, Inc., Carlsbad, CA, USA) at 24 h post-injection of LPCE or LPF1. In brief, a 2 mL volume of TRIzol was added to a 15 mL tube containing the homogenized frozen tissue of larvae and incubated at room temperature (RT) for 10 min. Subsequently, 400 μL of chloroform (Sigma-Aldrich, St. Louis, MO, USA) was added and the tubes were centrifuged at 12,000 × g for 15 min at 4°C. The supernatant was then transferred to a new tube, and 1 mL of isopropanol (Sigma-Aldrich, St. Louis, MO, USA) was added. After centrifugation, the obtained pellet was washed with 70% ethanol (Sigma-Aldrich, St. Louis, MO, USA), centrifuged again, and suspended in 50 μL of nuclease-free water (Ambion Inc., Carlsbad, CA, USA). The concentration, purity and quality of the RNA were verified using a NanoVue Plus spectrophotometer (GE Healthcare Bio-Sciences, Pittsburgh, PA, USA).
The extracted total RNA (1 μg) was transcribed into complementary DNA (cDNA) using the Verso cDNA Synthesis Kit (Thermo Fisher Scientific Inc, Waltham, MA, USA), according to the protocols recommended by the manufacturer. The primers for the genes that encode β-actin and galiomicin were designed by Rossoni et al. (2017) and described in Table 2. The transcribed cDNAs were amplified for relative quantification of the expression of the gene encoding galiomicin in relation to the concentration of the reference gene (β-actin).
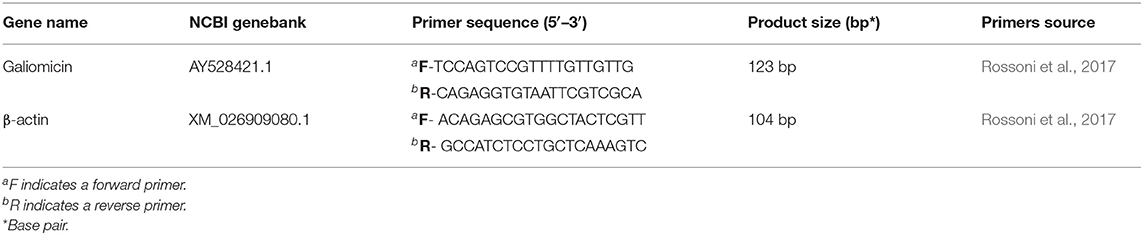
Table 2. List and description of genes and primers sequences used in the qPCR.
Statistical Analysis
The Student's t-test was used to compare the results from the cell-cell interaction, time to kill assay, and in vitro biofilm assay. Percent survival and killing curves of G. mellonella were plotted and statistical analysis was performed by the Kaplan-Meier test using Stata Statistical Software (Stata Corp LP, College Station, TX, USA). Analysis of variance (ANOVA) and Tukey post-test were used to compare the results obtained in the data of hemocyte count and in the analysis of gene expression. All the tests were performed using GraphPad Prism statistical software (GraphPad Software, Inc., California, CA, USA) and a P-value ≤ 0.05 was considered significant.
Results
Probiotic Effect
First, L. paracasei 28.4 was screened for antifungal activity against C. auris CAU-01 using co-culture. For this purpose, L. paracasei 28.4 cells were co-cultured with C. auris for 1–3 days. There was a significant reduction in C. auris counts for all evaluated times (1 day: 3.6 Log, 2 days: 1.67 Log, and 3 days: 1.8 Log) compared to the control group as demonstrated in Figure 1. L. paracasei concentration was constant throughout the assay, however C. auris experienced growth suppression in the presence of L. paracasei 28.4. This probiotic effect can indicate a better ability of L. paracasei to utilize nutrients in the media for growth, a direct cell-cell interaction, or possibly inhibition by bacterial metabolites.
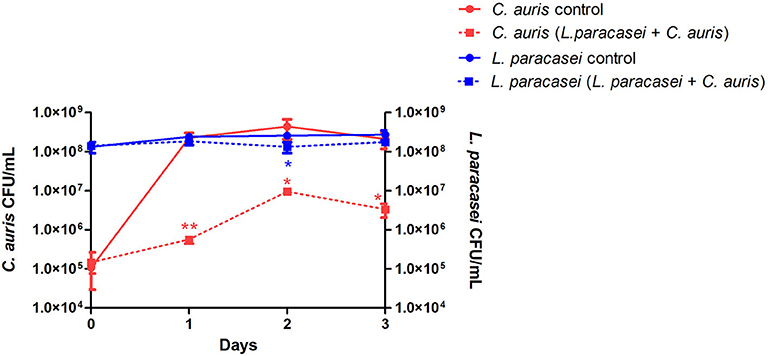
Figure 1. The viability of C. auris CAU-01 (solid line) was significantly reduced when co-cultured with L. paracasei 28.4 (dashed line). Student t-test, *p ≤ 0.01, **p ≤ 0.001.
Postbiotic Planktonic Inhibition
To explore the possibility of an indirect inhibitory activity posed by bacterial metabolites, we investigated C. auris inhibition by cell-free supernatant of L. paracasei 28.4. Inhibitory assessment was made using crude extract (LPCE) and a fraction (LPF1) derived from the L. paracasei supernatant. The MIC of LPCE and LPF1 was evaluated for 10 strains of C. auris (Table 3). MICs for LPCE were 15 mg/mL for all strains, and LPF1 MICs ranged from 3.75 to 7.5 mg/mL. For all strains evaluated, the MFC values were ≤ 4 × MIC, so the postbiotic elements were considered cidal (Figure 2). Thus, there does appear to be a postbiotic effect by L. paracasei 28.4. The lower MIC of LPCE is reasonable considering that the active component is more diluted in the unfractionated volume.
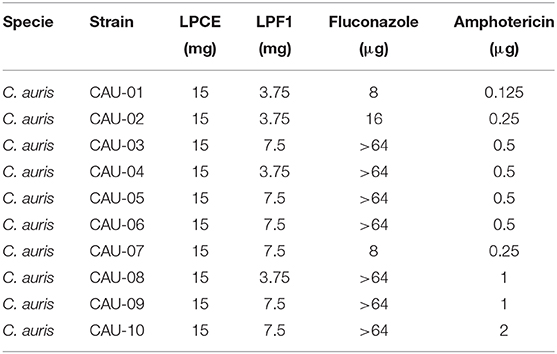
Table 3. MIC for C. auris strains.
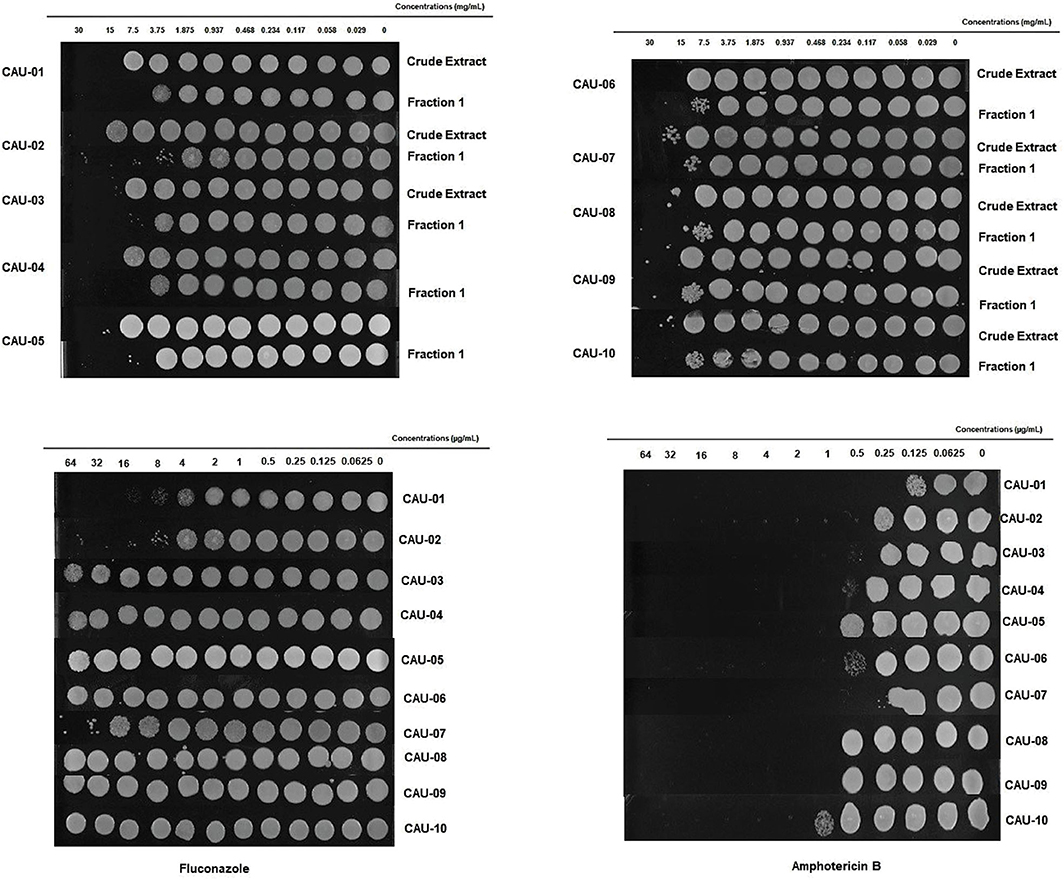
Figure 2. LPCE and LPF1 inhibits C. auris. The static vs. cidal nature of the inhibition was examined for the various postbiotics concentrations. Growth at concentration equivalent or higher than the MIC indicated that LPCE and LPF1 were fungistatic and inhibition of growth indicated fungicidal activity.
Since LPCE and LPF1 were active against all of the tested C. auris strains, we selected a single strain (CAU-01) for follow-up experiments. The killing kinetics assay determined that the total viable fungal count was about 6-log CFU at 0 h. After 24 h, there was a complete reduction of the total viable count of C. auris cells treated with LPCE (4 × MIC: 60 mg/mL) or LPF1 (8 × MIC: 30 mg/mL) (Figure 3). As a positive control, we included the antifungal agent fluconazole. Fluconazole decreased the initial inoculum by 1-log CFU during the same time period at all concentrations tested (1 × MIC: 4 μg/mL; 4 × MIC: 16 μg/mL, and 8 × MIC: 32 μg/mL).
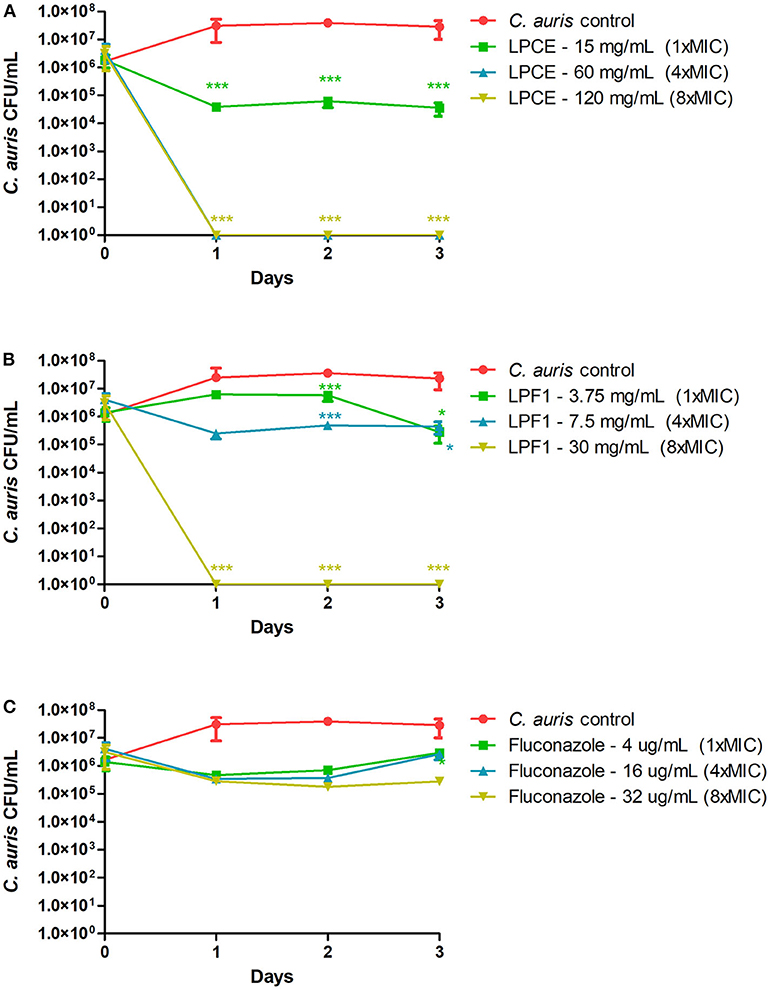
Figure 3. Killing kinetics. (A) Growth curves were generated using C. auris cells treated with LPCE. (B) Growth curves were generated using C. auris cells treated with LPF1. (C) Growth curves were generated using C. auris cells treated with fluconazole. Student t-test, *p ≤ 0.01, ***p ≤ 0.0001.
Postbiotic Biofilm Inhibition
It is known that antifungal compounds have variable efficacy against biofilms. Therefore, the cell free supernatant extract and fraction were tested for inhibitory activity on biofilms. LPCE and LPF1 at 1 × MIC and 2 × MIC concentrations significantly reduced the biomass (p = 0.0001) and the metabolic activity (p = 0.0001) of the C. auris biofilm as shown in Figures 4A,B, respectively. A biomass of C. auris showed a reduction of up to 67% for LPF1, 61% for LPCE, and 21% using fluconazole. The metabolic activity of biofilms reduced 89, 85, and 23% for LPF1, LPCE, and fluconazole, respectively. The biofilms treated with the postbiotic elements in the concentration of 0.5 × MIC also caused a reduction in relation to the control groups but there was no statistically significant difference, indicating a dose dependent effect on the biofilms.
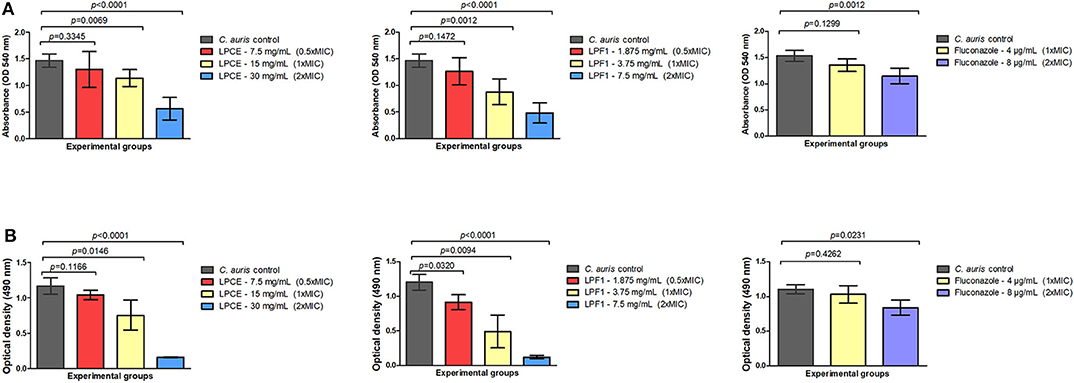
Figure 4. Quantification of cells in biofilms formed at the bottom of 96-well plates. (A) Biomass quantification and (B) Metabolic activity (XTT analysis) of different treatments (LPCE, LPF1, and Fluconazole). The different concentrations of LPCE and LPF1 used in biofilm formation correspond to 0.5 × MIC, 1 × MIC, and 2 × MIC values. Student's t-test, p ≤ 0.05.
We sought to determine the susceptibility of persister cells to the L. paracasei 28.4 supernatant derived elements. To generate C. auris persisters, C. auris CAU-01 was grown to stationary phase and then was treated with different concentrations of amphotericin and fluconazole (10x, 50x, 100x, 150x, and 200x MIC) or postbiotic elements (10 × MIC) for 48 h. The concentration of cells in stationary phase was 5 × 108 CFU/mL (~8.54 Log) and, after treating with 200 × MIC (25 μg/mL) amphotericin B or 200 × MIC (1,600 μg/mL) fluconazole for 48 h, the cell viability was ~104 CFU/mL and ~107 CFU/mL, respectively. Also, we treated the cells in stationary phase with a dose of 10 × MIC LPCE (150 mg/mL) or LPF1 (37.5 mg/mL) for 48 h and there was a complete reduction in the growth of C. auris cells. C. auris was tolerant of standard of care medications fluconazole and amphotericin at concentrations that are normally detrimental to the fungi, suggesting a persistent state. Under the same conditions, exposure to LPCE, and LPF elicited a cell reduction, suggesting the ability to inhibit persister cells (Figure 5).
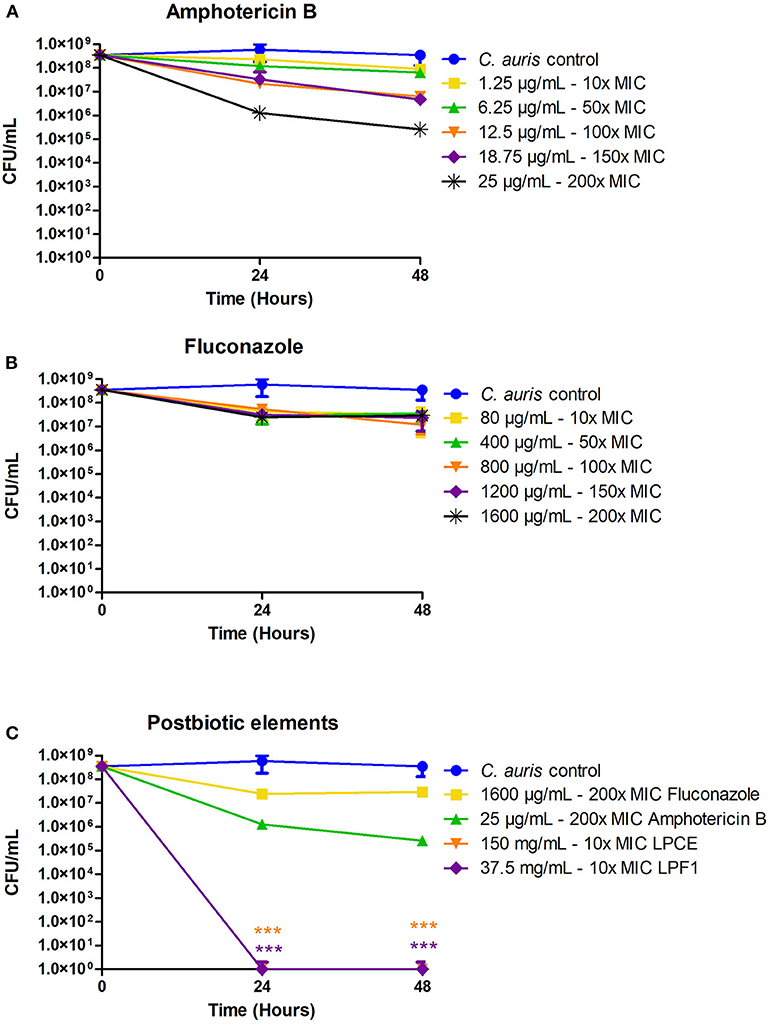
Figure 5. Isolation of C. auris persisters. (A) C. auris persisters cells induced by amphotericin B. (B) C. auris persisters cells induced by fluconazole. (C) LPCE and LPF1 were able to kill all persister cells. Student's t-test, ***p ≤ 0.0001.
Postbiotic Treatment in the G. mellonella Infection Model
First, G. mellonella was used to evaluate acute systemic toxicity of the postbiotic elements. The larvae were injected with varying concentrations of LPCE and LPF1 (80–10 mg/kg), and their survival was monitored for a period of 7 days. LPCE and LPF1 did not exert toxic effects on the larvae when administered at those concentrations (Figures 6A,B).
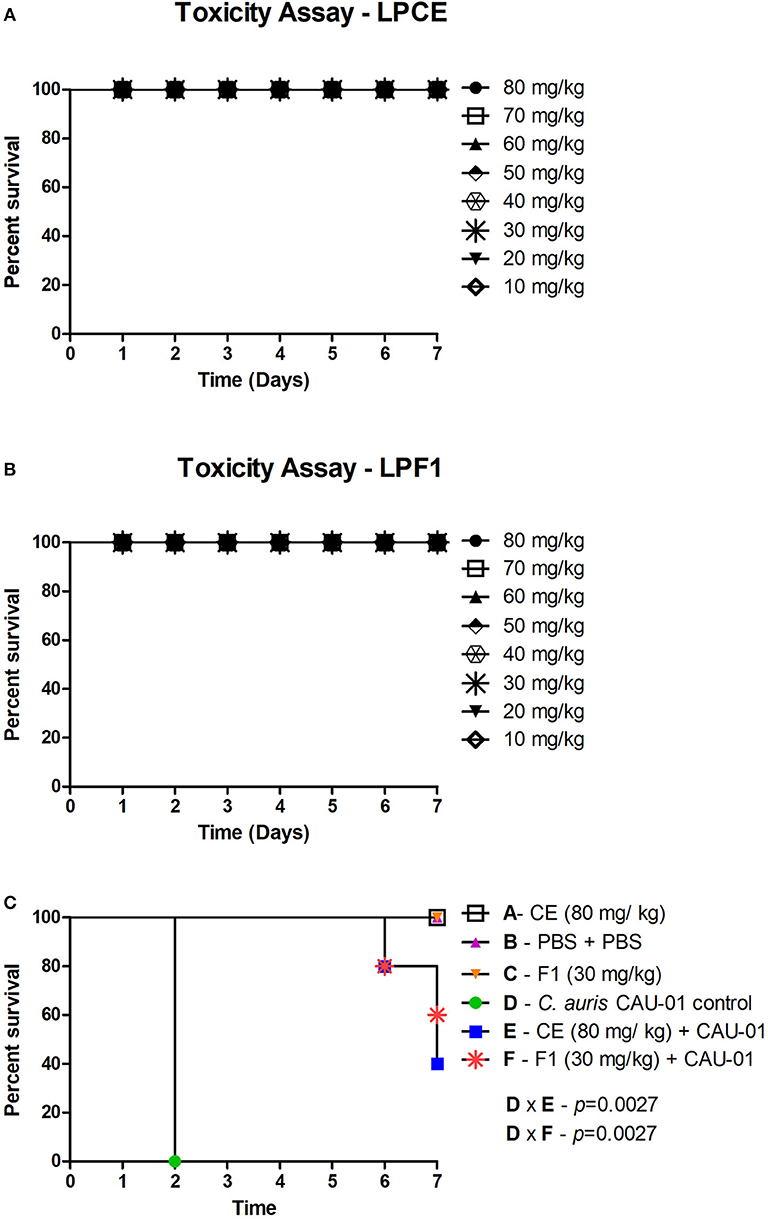
Figure 6. (A) Toxicity evaluation of LPCE in G. mellonella model. (B) Toxicity evaluation of LPF1 in G. mellonella model. G. mellonella larvae were injected with serial concentrations of postbiotic elements. No death was observed at the concentrations used. (C) LPCE and LPF1 prolong the survival of G. mellonella larvae infected with C. auris. Significant differences were observed in survival between the “LPCE (80 mg/mL) + CAU-01 group” and “PBS + C. auris CAU-01 control group” and between the “LPF1 (30 mg/mL) + CAU-01 group” and “PBS + C. auris CAU-01 control group.” Kaplan-Meier test, p ≤ 0.05.
To investigate the antifungal effects of LPF1 and LPCE in a G. mellonella model, we tested the efficacy of pretreatment with LPF1 and LPCE in larvae infected with C. auris. LPF1 and LPCE were injected into the larvae at 2 h prior to infection with C. auris concentrations of 30 and 80 mg/kg, respectively. In the control group, infection with C. auris without previous injection of postbiotics elements caused death in 100% of the larvae within 3 days. When the larvae were pretreated with LPF1 or LPCE prior to C. auris infection, the survival rate of G. mellonella larvae was significantly prolonged (p < 0.05) (Figure 6C). More specifically, larval survival increased 43% for LPF1 and 37% for LPCE groups.
To investigate the immune mechanisms associated with the effects of LPCE and LPF1 against C. auris infection, we determined the number of available hemocytes in the hemolymph of larvae after 4 h of C. auris injection. Hemocyte count was performed after 4 h of infection based on our previous study in which L. paracasei 28.4 stimulated hemocyte production in 4 h rather than 24 h (Rossoni et al., 2017). We analyzed only the larvae not infected by C. auris and observed an increase in the number of hemocyte in the LPCE (1.43-fold increase) (Figure 7A) and LPF1 (1.8-fold increase) (Figure 7B) groups compared to the PBS control group, but there was no statistically significant difference. In the larvae infected with C. auris, the groups pretreated with LPCE or LPF1 also showed increased hemocyte numbers compared to the C. auris control group (p < 0.05). The LPCE and LPFI groups reached, respectively, 2.55 (Figure 7A) and 2.26-fold increase (Figure 7B). Interestingly, we also observed that the C. auris group showed a reduction of hemocyte numbers in relation to the PBS control group in agreement with Bergin et al. (2003) that demonstrated an inverse relationship with infectious fungi and hemocyte density, but when the larvae were pretreated with LPCE or LPF1, the hemocyte quantity was very similar to the values found in the PBS control group (Figures 7A,B).
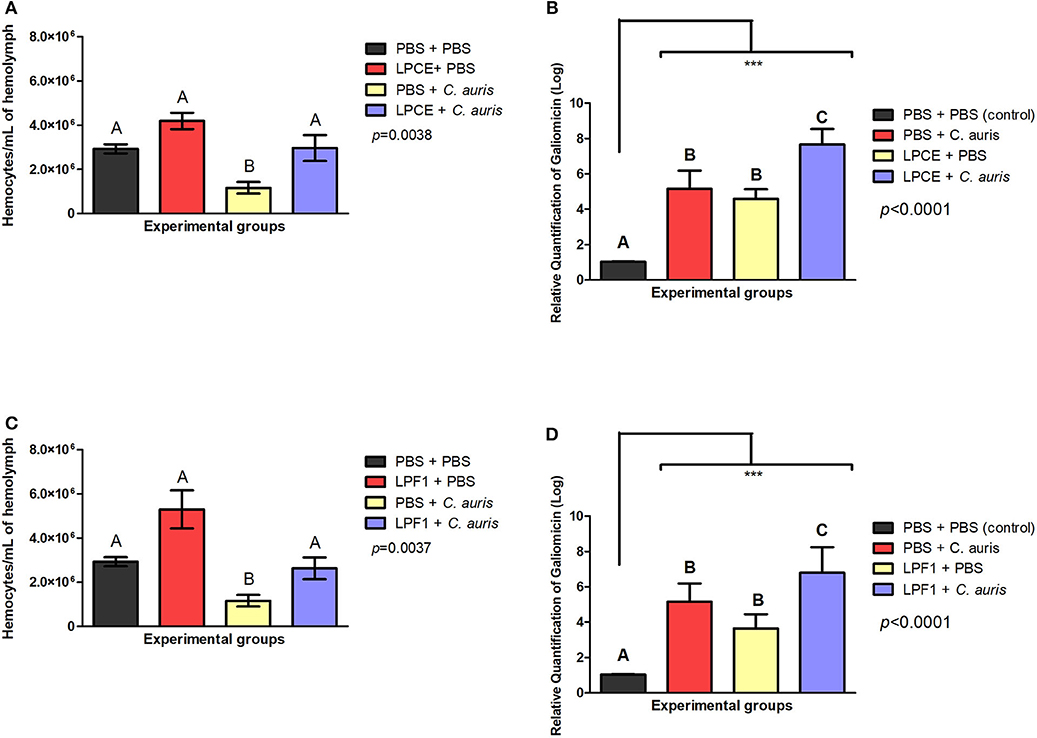
Figure 7. LPCE and LPF1 modulate the immune system of G. mellonella. (A) LPCE: The group of LPCE + PBS increased the hemocyte number compared to a PBS control (PBS + PBS). The group LPCE + C. auris also increased the hemocyte quantity compared to C. auris group (PBS + C. auris). (B) LPF1: The group of LPF1 + PBS increased the hemocyte number compared to a PBS control (PBS + PBS). The group LPF1 + C. auris also increased the hemocyte quantity compared to C. auris group (PBS + C. auris). PBS + C. auris group showed a reduction of hemocyte quantity in relation to the PBS control group, but when the larvae were pretreated with LPCE or LPF1, the hemocyte quantity was very similar to the values found in the PBS control group. (C,D) LPCE and LPF1 increased the gene expression of galiomicin of G. mellonella. Relative quantification (log) of galiomicin for the groups treated with only PBS (Control), pre-treated with PBS and infected with C. auris, only treated with LPCE or LPF1, and pre-treated with LPCE or LPF1 and infected with C. auris. The units in the Y-axis were calculated based on the 2−ΔΔCT method, and they are expressed as the means and standard deviation. Galiomicin expression was normalized and compared with the expression of insects exposed to the control (PBS) using the reference gene β-actin. Different letters (A, B, and C) represent statistically significant differences among the groups. ANOVA and Tukey Tests (p < 0.05 was considered significant). ***p ≤ 0.001.
The presence of an increased hemocyte count suggests that LPCE and LPF1 could modulate the immune response of G. mellonella larvae. Thus, we further explored alterations in the immune response examining the expression of the antifungal peptide galiomicin. For this assay, we evaluated the galiomicin expression after 24 h since the greatest expression of this peptide occurs at a later stage of Candida infection (Rossoni et al., 2017). We found that LPCE and LPF1 were able to increase the expression of galiomicin. The groups pretreated with LPCE or LPF1 and then infected with C. auris had a statistically significant increase (p < 0.0001) in relation to the control group infected by C. auris (LPCE group: 1.48-fold increase; LPF1 group: 1.31-fold increase). LPCE and LPF1 induced an increase in gene expression of 4.58 and 3.64-fold compared, respectively, to the control group formed by consecutive PBS injections (Figures 7C,D). These results indicate that LPCE and LPF1 increased the hemocyte density and levels of galiomicin expression, which may protect G. mellonella from C. auris infection.
Discussion
Recently, postbiotics have gained more and more attention due to their beneficial actions on the host without the adverse risk of inducing bacteremia in immunocompromised patients from the delivery of live cells (Gao et al., 2019). In this study, we identified LPCE and LPF1 derived from L. paracasei 28.4 supernatant as postbiotics and a potentially alternative antifungal treatment against C. auris, a globally emerging pathogen. The results obtained in the cell-cell interaction demonstrated that L. paracasei strain 28.4 was able to interfere with C. auris growth, demonstrating the potential for probiotic activity against C. auris. Also, the postbiotic elements reduced C. auris in planktonic, biofilm, and persister states, a significant feat. In in vivo assays, LPCE and LPF1 protected G. mellonella infected with C. auris. Our research provides a novel idea for prevention and treatment of C. auris infections.
The MIC results of this study demonstrated the antifungal activity of LPCE and LPF1 against all 10 C. auris strains, including both fluconazole-sensitive and fluconazole-resistant strains. Although the MIC values found were higher than fluconazole, the postbiotic elements are probably composed of a pool of molecules and are not yet fully purified. In addition, it was observed that the higher its purification and subsequent fractionation, the lower the MIC value obtained (MIC value of LPF1 is four times less than the value of LPCE).
The antibacterial properties of postbiotics have been tested on bacterial diseases; for example, Dunand et al. (2019) determined the protective capacity of postbiotics of fermented milk against Salmonella enterica serovar Typhimurium. The authors obtained the postbiotic from five frozen commercial cultures of thermophilic lactobacilli and 11 autochthonous Lactobacillus strains. The use of postbiotics for 14 days significantly increased the secretory IgA in feces of mice and a higher survival was also observed compared to controls demonstrating an immunomodulatory and protective capacity against S. enterica serovar Typhimurium infection in mice. In addition, studies demonstrate the beneficial role of postbiotics in inflammatory activity (Tsilingiri et al., 2012; Compare et al., 2017; Gao et al., 2019; Johnson et al., 2019). To our knowledge, this is the first study supporting the use of a postbiotic from Lactobacillus cells against C. auris and its use remains unexplored as a therapeutic option in these fungal infections. Therefore, further studies are needed to evaluate some important aspects such as toxicity, adverse effects, and viability of mass production.
To investigate the antifungal potential of LPCE and LPF1, we evaluated its effects in the growth kinetics of C. auris. The use of LPCE and LPF1 completely inhibited C. auris growth in contrast to the clinical antifungal agent fluconazole, which did not demonstrate any efficacy more than 1-Log. LPCE and LPF1 also exerted anti-biofilm activity against C. auris; significant reductions in biofilm formation were observed in both biomass amount and metabolic activity. In agreement to this result, Rossoni et al. (2018) evaluated the antifungal action of the L. paracasei 28.4 supernatant on different C. albicans strains. The raw supernatant of L. paracasei 28.4 was capable of reducing the growth of C. albicans by up to 73% in planktonic cultures, 62% in biofilms and interferes negatively in adhesion (ALS3: 66-fold decrease) and hyphae formation genes (HWP1: 66-fold decrease; CPH1: 1000-fold decrease). Although the postbiotic elements have had effectiveness to reduce C. auris biofilms, previous studies have shown that biofilm formation is highly variable between different strains of C. auris and this fact merits further exploration (Sherry et al., 2017; Pathirana et al., 2018).
The biofilms of C. auris may become increasingly resistant to conventional antifungals according to their formation time. For example, after 4 h of biofilm development, the median MIC increased 16-fold for miconazole and 4-fold for amphotericin B compared to the 0 h time of biofilm formation (Kean et al., 2018). Borman et al. (2016) demonstrated that old cultures of C. auris can survive in high concentrations of fluconazole (256 μg/mL), as well as be unresponsive to treatment in vivo using G. mellonella model. These facts agree with the low sensitivity of biofilms to fluconazole found in this study and reinforce the search for alternative treatments such as postbiotics in C. auris infections.
Biofilms harbor drug resistant cells, included among them are persister cells which, in their metabolically dormant state, can be recalcitrant to antifungal agents (LaFleur et al., 2006). One important aspect of postbiotic elements was their ability to eliminate all C. auris persister cells that survived high dosages of amphotericin B or fluconazole. Persister cells were reported for the first time as a subpopulation of bacteria tolerant to a particular antibiotic that, after removal of the antimicrobial agent, gave rise to a new population of susceptible microbial cells (Bigger, 1944). In the clinical setting, persisters are usually associated with recurrent infections and the development of chronic infections (Denega et al., 2019). The first report of Candida spp. persister cells was described for LaFleur et al. (2006) that showed a biphasic killing curve when C. albicans was exposed to amphotericin B, chlorhexidine, or a combination of both. In addition, Al-Dhaheri and Douglas (2008) showed that not all Candida spp. and strains are able to form persister cells, for example, C. albicans strain SC5314 (Gillum et al., 1984), one of the most commonly used C. albicans strains used for molecular genetics studies, is not able to form persister cells in vitro (Denega et al., 2019).
The alternative invertebrate model of G. mellonella was used to evaluate protective effects of LPCE and LPF1 in experimental candidiasis by C. auris. First, in order to evaluate acute systemic toxicity of the postbiotic elements, the larvae were injected with different doses of LPCE and LPF1, and none of the systemic doses (10–80 mg/kg) resulted in death of the larvae. Additionally, we found that the injection of LPCE or LPF1 into G. mellonella larvae infected by C. auris increased the survival of these insects (43% for LPF1; 37% for LPCE), the number of hemocytes and the gene expression of galiomicin. These data further confirm the excellent performance of LPCE and LPF1, thus providing important insight into combating C. auris. To the best of our knowledge, this is the first article in the literature that uses postbiotic elements in G. mellonella.
Taken together, these findings indicate that LPCE and LPF1 are capable of stimulating the cellular and humoral immune responses of the larvae and consequently reduce C. auris infection. The use of posbiotics elements with antifungal and immunomodulatory properties could be promising in C. auris infection, once a recent study questioned the effectiveness of the immune system against this species. It was demonstrated that human neutrophils do not properly recognize C. auris as they do with other Candida species and this behavior may explain the high mortality rates even for patients treated with antifungals (Nett, 2019).
Modulation of the G. mellonella immune response has been reported by different studies using probiotics (Ribeiro et al., 2017; Rossoni et al., 2017; Scalfaro et al., 2017; Geraldo et al., 2019). The main cells involved in the cellular immune response of G. mellonella are hemocytes (Bergin et al., 2003; Sheehan and Kavanagh, 2018). These are responsible for important events such as nodulation, encapsulation, and phagocytosis. Phagocytosis is a very important cellular process in which some enzymes are released by hemocytes in order to destroy the invading pathogen (Kavanagh and Reeves, 2004; Pereira et al., 2018; Sheehan and Kavanagh, 2018). The humoral response of these insects is constituted by effector molecules including opsonins, melanin, and antimicrobial peptides (AMPs) (Tsai et al., 2016). Although there are 18 different types of AMPs identified in G. mellonella hemolymph, we chose to examine galiomicin because it is a specific G. mellonella defense and is one of the most effective AMPs against fungal infection (Wojda, 2017). This AMP shows homology to human cysteine-rich peptides from the ß group of defensins and make up part of the innate immune system. In general, AMPs are the last line of defense and they act on the hemolymph attacking elements of the bacterial or fungal cell wall (Shai, 2002; Mowlds et al., 2010; Rossoni et al., 2017).
Although postbiotic elements of L. paracasei 28.4 have shown promising results, the key components of LPCE and LPF1 with antifungal properties are still unknown and need to be further investigated. Future studies should be addressed for the isolation and characterization of the bioactive molecules presents in L. paracasei 28.4 supernatant, as well as for the action mechanisms of these postbiotic elements on specific targets in fungal cells.
Within the limitations of the study, it can be concluded that L. paracasei 28.4 cells and its postbiotic elements (LPCE and LPF1) have antifungal activity against C. auris including activity against planktonic and persister cells, as well as biofilms. Postbiotic derived from L. paracasei 28.4 protected G. mellonella infected with C. auris and enhanced its immune status indicating a dual function in modulating the host immune response. The exact mechanisms related to the action of postbiotic elements against C. auris are still unclear and need to be further investigated.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Author Contributions
RR and PB conducted most of the experiments. DS, IM, and RM played a key role in extraction and fractionation of post-biotic elements. RR, BF, JJ, and EM wrote the paper. EM, BF, and JJ conceived and supervised all the work. All authors contributed to the article and approved the submitted version.
Funding
JJ was supported by the National Council for Scientific Development/CNPq (306330/2018-0). RR received a scholarship from São Paulo Research Foundation/FAPESP (Grants 2017/19219-3 and 2018/21239-5). BF was supported by NIH grant P20 GM121344.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
BHI, Brain heart infusion; CDC, Centers for Disease Control and Prevention; CFU, colony-forming unit; CLSI, Clinical and Laboratory Standards Institute; LPCE, L. paracasei crude extract; LPF1, L. paracasei fraction 1; MIC, minimum inhibitory concentration; MRS, Man, Rogosa and Sharpe; PBS, phosphate buffered saline; YNB, Yeast nitrogen base; YPD, Yeast peptone dextrose.
References
Al-Dhaheri, R. S., and Douglas, L. J. (2008). Absence of amphotericin B-tolerant persister cells in biofilms of some Candida species. Antimicrob. Agents Chemother. 52, 1884–1887. doi: 10.1128/AAC.01473-07
Bergin, D., Brennan, M., and Kavanagh, K. (2003). Fluctuations in haemocyte density and microbial load may be used as indicators of fungal pathogenicity in larvae of Galleria mellonella. Microbes Infect. 5, 1389–1395. doi: 10.1016/j.micinf.2003.09.019
Bidaud, A. L., Chowdhary, A., and Dannaoui, E. (2018). Candida auris: an emerging drug resistant yeast—a mini-review. J. Mycol. Med. 28, 568–573. doi: 10.1016/j.mycmed.2018.06.007
Bigger, J. (1944). Treatment of Staphylococcal infections with penicillin by intermittent sterilisation. Lancet 244, 497–500. doi: 10.1016/S0140-6736(00)74210-3
Borman, A. M., Szekely, A., and Johnson, E. M. (2016). Comparative pathogenicity of United Kingdom isolates of the emerging pathogen Candida auris and other key pathogenic candida species. mSphere 1, e00189–e00216. doi: 10.1128/mSphere.00189-16
Cannon, J. P., Lee, T. A., Bolanos, J. T., and Danziger, L. H. (2005). Pathogenic relevance of Lactobacillus: a retrospective review of over 200 cases. Eur. J. Clin. Microbiol. Infect. Dis. 24, 31–40. doi: 10.1007/s10096-004-1253-y
Centers for Disease Control and Prevention (2016). Global Emergence of Invasive Infections Caused by the Multidrug-Resistant Yeast Candida auris. Atlanta, GA: Clinical Alert to US Healthcare Facilities.
Cernakova, L., Light, C., Salehi, B., Rogel-Castillo, C., Victoriano, M., Martorell, M., et al. (2019). Novel therapies for biofilm-based Candida spp. infections. Adv. Exp. Med. Biol. 1214, 93–123. doi: 10.1007/5584_2019_400
Chaabane, F., Graf, A., Jequier, L., and Coste, A. T. (2019). Review on antifungal resistance mechanisms in the emerging pathogen Candida auris. Front. Microbiol. 10:2788. doi: 10.3389/fmicb.2019.02788
Chakrabarti, A., and Singh, S. (2020). Multidrug-resistant Candida auris: an epidemiological review. Expert Rev. Anti Infect. Ther. 18, 551–562. doi: 10.1080/14787210.2020.1750368
Cicenia, A., Scirocco, A., Carabotti, M., Pallotta, L., Marignani, M., and Severi, C. (2014). Postbiotic activities of lactobacilli-derived factors. J. Clin. Gastroenterol. 48(Suppl.1), S18–S22. doi: 10.1097/MCG.0000000000000231
Compare, D., Rocco, A., Coccoli, P., Angrisani, D., Sgamato, C., Iovine, B., et al. (2017). Lactobacillus casei DG and its postbiotic reduce the inflammatory mucosal response: an ex-vivo organ culture model of post-infectious irritable bowel syndrome. BMC Gastroenterol. 17:53. doi: 10.1186/s12876-017-0605-x
Davies, D. (2003). Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2, 114–122. doi: 10.1038/nrd1008
de Barros, P. P., Scorzoni, L., Ribeiro, F. C., Fugisaki, L. R. O., Fuchs, B. B., Mylonakis, E., et al. (2018). Lactobacillus paracasei 28.4 reduces in vitro hyphae formation of Candida albicans and prevents the filamentation in an experimental model of Caenorhabditis elegans. Microb. Pathog. 117, 80–87. doi: 10.1016/j.micpath.2018.02.019
Denega, I., d'Enfert, C., and Bachellier-Bassi, S. (2019). Candida albicans biofilms are generally devoid of persister cells. Antimicrob. Agents Chemother. 63:e01979-18. doi: 10.1128/AAC.01979-18
Dunand, E., Burns, P., Binetti, A., Bergamini, C., Peralta, G. H., Forzani, L., et al. (2019). Postbiotics produced at laboratory and industrial level as potential functional food ingredients with the capacity to protect mice against Salmonella infection. J. Appl. Microbiol. 127, 219–229. doi: 10.1111/jam.14276
Gao, J., Li, Y., Wan, Y., Hu, T., Liu, L., Yang, S., et al. (2019). A novel postbiotic from Lactobacillus rhamnosus GG with a beneficial effect on intestinal barrier function. Front. Microbiol. 10:477. doi: 10.3389/fmicb.2019.00477
Geraldo, B. M. C., Batalha, M. N., Milhan, N. V. M., Rossoni, R. D., Scorzoni, L., and Anbinder, A. L. (2019). Heat-killed Lactobacillus reuteri and cell-free culture supernatant have similar effects to viable probiotics during interaction with Porphyromonas gingivalis. J. Periodontal. Res. 55, 215–220. doi: 10.1111/jre.12704
Gillum, A. M., Tsay, E. Y., and Kirsch, D. R. (1984). Isolation of the Candida albicans gene for orotidine-5'-phosphate decarboxylase by complementation of S. cerevisiae ura3 and E. coli pyrF mutations. Mol. Gen. Genet. 198, 179–182. doi: 10.1007/BF00328721
Guarner, F., Khan, A. G., Garisch, J., Eliakim, R., Gangl, A., Thomson, A., et al. (2012). World gastroenterology organisation global guidelines: probiotics and prebiotics october 2011. J. Clin. Gastroenterol. 46, 468–481. doi: 10.1097/MCG.0b013e3182549092
Hu, H. J., Zhang, G. Q., Zhang, Q., Shakya, S., and Li, Z. Y. (2017). Probiotics prevent Candida colonization and invasive fungal sepsis in preterm neonates: a systematic review and meta-analysis of randomized controlled trials. Pediatr. Neonatol. 58, 103–110. doi: 10.1016/j.pedneo.2016.06.001
Jacobsen, I. D., Wilson, D., Wachtler, B., Brunke, S., Naglik, J. R., and Hube, B. (2012). Candida albicans dimorphism as a therapeutic target. Expert Rev. Anti Infect. Ther. 10, 85–93. doi: 10.1586/eri.11.152
Janczarek, M., Bachanek, T., Mazur, E., and Chalas, R. (2016). The role of probiotics in prevention of oral diseases. Postepy Hig. Med. Dosw. 70, 850–857. doi: 10.5604/17322693.1214381
Jin, Y., Samaranayake, L. P., Samaranayake, Y., and Yip, H. K. (2004). Biofilm formation of Candida albicans is variably affected by saliva and dietary sugars. Arch. Oral Biol. 49, 789–798. doi: 10.1016/j.archoralbio.2004.04.011
Johnson, C. N., Kogut, M. H., Genovese, K., He, H., Kazemi, S., and Arsenault, R. J. (2019). Administration of a postbiotic causes immunomodulatory responses in broiler gut and reduces disease pathogenesis following challenge. Microorganisms 7:268. doi: 10.3390/microorganisms7080268
Kavanagh, K., and Reeves, E. P. (2004). Exploiting the potential of insects for in vivo pathogenicity testing of microbial pathogens. FEMS Microbiol. Rev. 28, 101–112. doi: 10.1016/j.femsre.2003.09.002
Kean, R., Delaney, C., Sherry, L., Borman, A., Johnson, E. M., Richardson, M. D., et al. (2018). Transcriptome assembly and profiling of Candida auris reveals novel insights into biofilm-mediated resistance. mSphere 3, e00334–e00418. doi: 10.1128/mSphere.00334-18
LaFleur, M. D., Kumamoto, C. A., and Lewis, K. (2006). Candida albicans biofilms produce antifungal-tolerant persister cells. Antimicrob. Agents Chemother. 50, 3839–3846. doi: 10.1128/AAC.00684-06
Lockhart, S. R., Etienne, K. A., Vallabhaneni, S., Farooqi, J., Chowdhary, A., Govender, N. P., et al. (2017). Simultaneous emergence of multidrug-resistant Candida auris on 3 continents confirmed by whole-genome sequencing and epidemiological analyses. Clin. Infect. Dis. 64, 134–140. doi: 10.1093/cid/ciw691
Mailander-Sanchez, D., Wagener, J., and Schaller, M. (2012). Potential role of probiotic bacteria in the treatment and prevention of localised candidosis. Mycoses 55, 17–26. doi: 10.1111/j.1439-0507.2010.01967.x
Martins, N., Ferreira, I. C., Barros, L., Silva, S., and Henriques, M. (2014). Candidiasis: predisposing factors, prevention, diagnosis and alternative treatment. Mycopathologia 177, 223–240. doi: 10.1007/s11046-014-9749-1
Matsubara, V. H., Bandara, H. M., Mayer, M. P., and Samaranayake, L. P. (2016a). Probiotics as antifungals in mucosal candidiasis. Clin. Infect. Dis. 62, 1143–1153. doi: 10.1093/cid/ciw038
Matsubara, V. H., Wang, Y., Bandara, H. M., Mayer, M. P., and Samaranayake, L. P. (2016b). Probiotic lactobacilli inhibit early stages of Candida albicans biofilm development by reducing their growth, cell adhesion, and filamentation. Appl. Microbiol. Biotechnol. 100, 6415–6426. doi: 10.1007/s00253-016-7527-3
Medina, R. P., Araujo, A. R., Batista, J. M. Jr., Cardoso, C. L., Seidl, C., Vilela, A. F. L., et al. (2019). Botryane terpenoids produced by Nemania bipapillata, an endophytic fungus isolated from red alga Asparagopsis taxiformis—Falkenbergia stage. Sci. Rep. 9:12318. doi: 10.1038/s41598-019-48655-7
Mosca, F., Gianni, M. L., and Rescigno, M. (2019). Can postbiotics represent a new strategy for NEC? Adv. Exp. Med. Biol. 1125, 37–45. doi: 10.1007/5584_2018_314
Mowlds, P., Coates, C., Renwick, J., and Kavanagh, K. (2010). Dose-dependent cellular and humoral responses in Galleria mellonella larvae following beta-glucan inoculation. Microbes Infect. 12, 146–153. doi: 10.1016/j.micinf.2009.11.004
Mylonakis, E., Moreno, R., El Khoury, J. B., Idnurm, A., Heitman, J., Calderwood, S. B., et al. (2005). Galleria mellonella as a model system to study Cryptococcus neoformans pathogenesis. Infect. Immun. 73, 3842–3850. doi: 10.1128/IAI.73.7.3842-3850.2005
National Committee for Clinical Laboratory Standards (2002). “Reference method for broth dilution antifungal susceptibility testing of yeasts,” in M27-A Wayne (Pennsylvania, PA: Approved standard M27–A).
Nett, J. E. (2019). Candida auris: an emerging pathogen “incognito?” PLoS Pathog. 15:e1007638. doi: 10.1371/journal.ppat.1007638
Pathirana, R. U., Friedman, J., Norris, H. L., Salvatori, O., McCall, A. D., Kay, J., et al. (2018). Fluconazole-resistant Candida auris is susceptible to salivary histatin 5 killing and to intrinsic host defenses. Antimicrob. Agents Chemother. 62, e01872–e01917. doi: 10.1128/AAC.01872-17
Peeters, E., Nelis, H. J., and Coenye, T. (2008). Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 72, 157–165. doi: 10.1016/j.mimet.2007.11.010
Peleg, A. Y., Tampakakis, E., Fuchs, B. B., Eliopoulos, G. M., Moellering, R. C. Jr., and Mylonakis, E. (2008). Prokaryote-eukaryote interactions identified by using Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A. 105, 14585–14590. doi: 10.1073/pnas.0805048105
Pereira, T. C., de Barros, P. P., Fugisaki, L. R. O., Rossoni, R. D., Ribeiro, F. C., de Menezes, R. T., et al. (2018). Recent advances in the use of galleria mellonella model to study immune responses against human pathogens. J. Fungi. 4:128. doi: 10.3390/jof4040128
Pfaller, M. A., Sheehan, D. J., and Rex, J. H. (2004). Determination of fungicidal activities against yeasts and molds: lessons learned from bactericidal testing and the need for standardization. Clin. Microbiol. Rev. 17, 268–280. doi: 10.1128/CMR.17.2.268-280.2004
Ribeiro, F. C., de Barros, P. P., Rossoni, R. D., Junqueira, J. C., and Jorge, A. O. (2017). Lactobacillus rhamnosus inhibits Candida albicans virulence factors in vitro and modulates immune system in Galleria mellonella. J. Appl. Microbiol. 122, 201–211. doi: 10.1111/jam.13324
Ribeiro, F. C., Junqueira, J. C., Dos Santos, J. D., de Barros, P. P., Rossoni, R. D., Shukla, S., et al. (2020). Development of probiotic formulations for oral candidiasis prevention: Gellan gum as a carrier to deliver Lactobacillus paracasei 28.4. Antimicrob. Agents Chemother. 6, e02323–e02419. doi: 10.1128/AAC.02323-19
Riccardi, N., Rotulo, G. A., and Castagnola, E. (2019). Definition of opportunistic infections in immunocompromised children on the basis of etiologies and clinical features: a summary for practical purposes. Curr. Pediatr. Rev. 15, 197–206. doi: 10.2174/1573396315666190617151745
Rodrigues, C. F., Rodrigues, M. E., and Henriques, M. C. R. (2019). Promising alternative therapeutics for oral candidiasis. Curr. Med. Chem. 26, 2515–2528. doi: 10.2174/0929867325666180601102333
Rodrigues, M. E., Gomes, F., and Rodrigues, C. F. (2019). Candida spp./bacteria mixed biofilms. J. Fungi. 6:5. doi: 10.3390/jof6010005
Rodrigues, M. E., Silva, S., Azeredo, J., and Henriques, M. (2016). Novel strategies to fight Candida species infection. Crit. Rev. Microbiol. 42, 594–606. doi: 10.3109/1040841X.2014.974500
Rossato, L., and Colombo, A. L. (2018). Candida auris: what have we learned about its mechanisms of pathogenicity? Front. Microbiol. 9:3081. doi: 10.3389/fmicb.2018.03081
Rossoni, R. D., de Barros, P. P., de Alvarenga, J. A., Ribeiro, F. C., Velloso, M. D. S., Fuchs, B. B., et al. (2018). Antifungal activity of clinical Lactobacillus strains against Candida albicans biofilms: identification of potential probiotic candidates to prevent oral candidiasis. Biofouling 34, 212–225. doi: 10.1080/08927014.2018.1425402
Rossoni, R. D., de Barros, P. P., Lopes, L., Ribeiro, F. C., Nakatsuka, T., Kasaba, H., et al. (2019). Effects of surface pre-reacted glass-ionomer (S-PRG) eluate on Candida spp: antifungal activity, anti-biofilm properties, and protective effects on Galleria mellonella against C. albicans infection. Biofouling 35, 997–1006. doi: 10.1080/08927014.2019.1686485
Rossoni, R. D., Fuchs, B. B., Barros, P. P. d., Velloso M. d. S., Jorge, A. O. C., Junqueira, J. C., and Mylonakis, E. (2017). Lactobacillus paracasei modulates the immune system of Galleria mellonella and protects against Candida albicans infection. PLoS ONE 12:e0173332. doi: 10.1371/journal.pone.0173332
Sabino, R., Verissimo, C., Pereira, A. A., and Antunes, F. (2020). Candida auris, an agent of hospital-associated outbreaks: which challenging issues do we need to have in mind? Microorganisms 8:181. doi: 10.3390/microorganisms8020181
Salminen, M. K., Rautelin, H., Tynkkynen, S., Poussa, T., Saxelin, M., Valtonen, V., et al. (2006). Lactobacillus bacteremia, species identification, and antimicrobial susceptibility of 85 blood isolates. Clin. Infect. Dis. 42, e35–e44. doi: 10.1086/500214
Santos, R. B., Scorzoni, L., Namba, A. M., Rossoni, R. D., Jorge, A. O. C., and Junqueira, J. C. (2019). Lactobacillus species increase the survival of Galleria mellonella infected with Candida albicans and non-albicans Candida clinical isolates. Med. Mycol. 57, 391–394. doi: 10.1093/mmy/myy032
Scalfaro, C., Iacobino, A., Nardis, C., and Franciosa, G. (2017). Galleria mellonella as an in vivo model for assessing the protective activity of probiotics against gastrointestinal bacterial pathogens. FEMS Microbiol. Lett. 364, 1–6. doi: 10.1093/femsle/fnx064
Sekyere, J. O., and Asante, J. (2018). Emerging mechanisms of antimicrobial resistance in bacteria and fungi: advances in the era of genomics. Future Microbiol. 13, 241–262. doi: 10.2217/fmb-2017-0172
Serra, D. O., Klauck, G., and Hengge, R. (2015). Vertical stratification of matrix production is essential for physical integrity and architecture of macrocolony biofilms of Escherichia coli. Environ. Microbiol. 17, 5073–5088. doi: 10.1111/1462-2920.12991
Shai, Y. (2002). Mode of action of membrane active antimicrobial peptides. Biopolymers 66, 236–248. doi: 10.1002/bip.10260
Sheehan, G., and Kavanagh, K. (2018). Analysis of the early cellular and humoral responses of Galleria mellonella larvae to infection by Candida albicans. Virulence 9, 163–172. doi: 10.1080/21505594.2017.1370174
Sherry, L., Ramage, G., Kean, R., Borman, A., Johnson, E. M., Richardson, M. D., et al. (2017). Biofilm-forming capability of highly virulent, multidrug-resistant Candida auris. Emerg. Infect. Dis. 23, 328–331. doi: 10.3201/eid2302.161320
Spivak, E. S., and Hanson, K. E. (2018). Candida auris: an emerging fungal pathogen. J. Clin. Microbiol. 56, e01588–e01617. doi: 10.1128/JCM.01588-17
Tharmalingam, N., Khader, R., Fuchs, B. B., and Mylonakis, E. (2019). The anti-virulence efficacy of 4-(1,3-dimethyl-2,3-dihydro-1h-benzimidazol-2-yl)phenol against methicillin-resistant Staphylococcus aureus. Front. Microbiol. 10:1557. doi: 10.3389/fmicb.2019.01557
Tsai, C. J., Loh, J. M., and Proft, T. (2016). Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 7, 214–229. doi: 10.1080/21505594.2015.1135289
Tsilingiri, K., Barbosa, T., Penna, G., Caprioli, F., Sonzogni, A., Viale, G., et al. (2012). Probiotic and postbiotic activity in health and disease: comparison on a novel polarised ex-vivo organ culture model. Gut 61, 1007–1015. doi: 10.1136/gutjnl-2011-300971
Tsilingiri, K., and Rescigno, M. (2013). Postbiotics: what else? Benef. Microbes. 4, 101–107. doi: 10.3920/BM2012.0046
Vicariotto, F., Del Piano, M., Mogna, L., and Mogna, G. (2012). Effectiveness of the association of 2 probiotic strains formulated in a slow release vaginal product, in women affected by vulvovaginal candidiasis: a pilot study. J. Clin. Gastroenterol. 46, S73–S80. doi: 10.1097/MCG.0b013e3182684d71
Vilela, S. F., Barbosa, J. O., Rossoni, R. D., Santos, J. D., Prata, M. C., Anbinder, A. L., et al. (2015). Lactobacillus acidophilus ATCC 4356 inhibits biofilm formation by C. albicans and attenuates the experimental candidiasis in Galleria mellonella. Virulence 6, 29–39. doi: 10.4161/21505594.2014.981486
Wachtler, B., Citiulo, F., Jablonowski, N., Forster, S., Dalle, F., Schaller, M., et al. (2012). Candida albicans-epithelial interactions: dissecting the roles of active penetration, induced endocytosis and host factors on the infection process. PLoS ONE 7:e36952. doi: 10.1371/journal.pone.0036952
Wegh, C. A. M., Geerlings, S. Y., Knol, J., Roeselers, G., and Belzer, C. (2019). Postbiotics and their potential applications in early life nutrition and beyond. Int. J. Mol. Sci. 20:4673. doi: 10.3390/ijms20194673
Keywords: probiotic, postbiotic, Lactobacillus, Candida auris, biofilms
Citation: Rossoni RD, de Barros PP, Mendonça IC, Medina RP, Silva DHS, Fuchs BB, Junqueira JC and Mylonakis E (2020) The Postbiotic Activity of Lactobacillus paracasei 28.4 Against Candida auris. Front. Cell. Infect. Microbiol. 10:397. doi: 10.3389/fcimb.2020.00397
Received: 14 February 2020; Accepted: 29 June 2020;
Published: 04 August 2020.
Edited by:
Priya Uppuluri, University of California, Los Angeles, United StatesReviewed by:
Taissa Vila, University of Maryland, Baltimore, United StatesVishukumar Aimanianda, Institut Pasteur, France
Copyright © 2020 Rossoni, de Barros, Mendonça, Medina, Silva, Fuchs, Junqueira and Mylonakis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juliana Campos Junqueira, anVsaWFuYS5qdW5xdWVpcmFAdW5lc3AuYnI=