
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol., 20 May 2020
Sec. Clinical Microbiology
Volume 10 - 2020 | https://doi.org/10.3389/fcimb.2020.00200
 Courtney D. Petro1
Courtney D. Petro1 Jeffrey K. Duncan1
Jeffrey K. Duncan1 Yuliya I. Seldina2
Yuliya I. Seldina2 Anna Allué-Guardia3,4
Anna Allué-Guardia3,4 Mark Eppinger3,4Mark S. Riddle5David R. Tribble5Ryan C. Johnson5
Mark Eppinger3,4Mark S. Riddle5David R. Tribble5Ryan C. Johnson5 Clifton L. Dalgard6,7
Clifton L. Dalgard6,7 Gauthaman Sukumar7,8
Gauthaman Sukumar7,8 Patrick Connor9
Patrick Connor9 Nadia Boisen10
Nadia Boisen10 Angela R. Melton-Celsa1*
Angela R. Melton-Celsa1*To discern if there was a particular genotype associated with clinical enteroaggregative Escherichia coli (EAEC) strains isolated from deployed military personnel (DMP) with travelers' diarrhea (TD), we characterized a collection of EAEC from DMP deployed to Afghanistan, Djibouti, Kenya, or Honduras. Although we did not identify a specific EAEC genotype associated with TD in DMP, we found that EAEC isolated at the first clinic visit were more likely to encode the dispersin gene aap than EAEC collected at follow-up visits. A majority of the EAEC isolates were typical EAEC that adhered to HEp-2 cells, formed biofilms, and harbored genes for aggregative adherence fimbriae (AAF), AggR, and serine protease autotransporters of Enterobacteriaceae (SPATEs). A separate subset of the EAEC had aggR and genes for SPATEs but encoded a gene highly homologous to that for CS22, a fimbriae more commonly found in enterotoxigenic E. coli. None of these CS22-encoding EAEC formed biofilms in vitro or adhered to HEp-2 cells. Whole genome sequence and single nucleotide polymorphism analyses demonstrated that most of the strains were genetically diverse, but that a few were closely related. Isolation of these related strains occurred within days to more than a year apart, a finding that suggests a persistent source and genomic stability. In an ampicillin-treated mouse model we found that an agg4A+ aar- isolate formed a biofilm in the intestine and caused reduced weight gain in mice, whereas a strain that did not form an in vivo biofilm caused no morbidity. Our diverse strain collection from DMP displays the heterogeneity of EAEC strains isolated from human patients, and our mouse model of infection indicated the genotype agg4A+ aar– and/or capacity to form biofilm in vivo may correlate to disease severity.
Enteroaggregative Escherichia coli (EAEC) is a causative agent of both acute and persistent diarrhea in adults and children (Okeke and Nataro, 2001). Clinical presentation of EAEC infection is characterized by watery unformed stool with low-grade fever (Okeke and Nataro, 2001). For civilian travelers, EAEC is typically the third most identified enteropathogen responsible for TD, whereas EAEC is the second most common bacterial etiologic agent of TD in military travelers (Zboromyrska et al., 2014; Zaidi and Wine, 2015; Porter et al., 2017; Olson et al., 2019). For those deployed military personnel (DMP), TD is a leading cause of lost duty-days (Porter et al., 2017).
EAEC was originally characterized by the capacity to adhere to epithelial cells in a “stacked-brick” pattern (Nataro et al., 1987). Additionally, EAEC display aggregative adherence (AA) to one another and to synthetic surfaces, forming robust biofilms on plastic and glass (Okeke and Nataro, 2001), as well as in vivo in the ampicillin (Amp)-treated mouse model (Zangari et al., 2013). There are five different types of AA fimbriae (AAF), designated AAF/I-V, that are specific to EAEC (Jonsson et al., 2015). The genes for the AAF are located on the plasmid of aggregative adherence (pAA), and except for one reported strain (Jonsson et al., 2017), EAEC only encode one of the five AAF types. In the prototype strain 042 (AAF/II), proper display of fimbriae on the bacterial surface is dependent on the dispersin protein, Aap (Sheikh et al., 2002). Secretion of dispersin occurs through a dedicated transport system encoded on the pAA (genes aatPABCD). All of the genes necessary for AA are encoded on the pAA plasmid. Additional genes on pAA that are important for adherence include aggR, which encodes a transcriptional regulator. AggR positively regulates expression of genes required for AA as well as other genes on both pAA and the chromosome (Morin et al., 2013). Two examples of genes commonly found in EAEC that are regulated by AggR include the type 6 secretion system (T6SS) effector of unknown function, aaiC, and aatA, which encodes a component of the dispersin transport system. Besides AggR, many EAEC encode Aar, a negative regulator of AggR (Santiago et al., 2014). Mutations that inactivate aar cause increased fimbrial expression and elevated AggR expression (Santiago et al., 2014). Finally, a regulator encoded by eilA was shown in some EAEC to influence the capacity to adhere to HEp-2 cells and form biofilms (Sheikh et al., 2006). Other factors that may contribute to adherence by EAEC include heat-resistant hemagglutinin (Bhargava et al., 2009), long polar fimbriae (Ross et al., 2015), E. coli common pilus (Avelino et al., 2010), and Pil (Dudley et al., 2006; Garcia et al., 2019).
Once EAEC have established adherence, they can further damage epithelium through the release of a variety of factors that include members of the family of serine protease autotransporters of the Enterobacteriaceae (SPATEs). EAEC may encode one or several class 1 or 2 SPATEs (see review Ruiz-Perez and Nataro, 2014). Class 1 SPATEs that may be found in EAEC include the plasmid-encoded toxin Pet, which causes loss of actin stress fibers in tissue culture cells and subsequent cell death (Canizalez-Roman and Navarro-Garcia, 2003), Sat, which may affect tight junctions (Guignot et al., 2007), and SigA, which contributes to cell death and fluid accumulation in a rat ileal loop model of Shigella flexneri (Al-Hasani et al., 2000). Class 2 SPATEs generally exhibit immunomodulatory activity. For example, Pic exhibits proteolytic, mucinolytic activities and contributes to serum resistance (Harrington et al., 2009). SepA is a class 2 SPATE also found in S. flexneri where it has been found to contribute to disruption of the epithelial layer in model systems (Benjelloun-Touimi et al., 1995; Maldonado-Contreras et al., 2017). Classification and regression tree analysis (CART) showed that sepA from EAEC was associated more frequently with diarrheal cases than with controls in a study looking at pediatric cases of infection in Mali (Boisen et al., 2012). CART analysis was also used to examine EAEC strains in a case-control study in Brazil. In that study, the combination of pet and aafA (encodes AAF/II) was correlated with disease (Lima et al., 2013). A later study found that strains that lacked aar and pic were associated with malnourished children in Brazil (Havt et al., 2017).
In this investigation, we characterized a collection of EAEC strains isolated from DMP enrolled in the Trial Evaluating Ambulatory Therapy of Travelers' Diarrhea (TrEAT TD) Study (Riddle et al., 2017) by whole genome sequencing and analysis, biofilm formation, adherence to HEp-2 cells, and, for a subset, the capacity to cause failure-to-thrive (FTT) in an Amp-treated mouse model with the goal of identifying a genotype more associated with TD in the DMP population.
A collection of 174 presumptive EAEC (positive by PCR for aatA and/or aaiC) isolated from 98 patients during the TrEAT TD study (Riddle et al., 2017) were sent to Uniformed Services University of the Health Sciences. A secondary PCR screen of the isolates in our laboratory found that some of the isolates did not have either aatA or aaiC, and those isolates were excluded from the study. For our purposes, we named the cultures with a single letter, E (Egypt), K (Kenya), or P (Peru) to indicate the referral lab location in which the bacteria were isolated, followed by a number, then V1 (initial clinic visit), V4 (day 7 after initial clinic visit), or V5 (day 21 after initial clinic visit) to indicate the clinic visit from which the cultures arose. Isolates that were positive in our laboratory by PCR for aatA and/or aaiC were then screened by PCR for AAF and SPATE genes as well as for aggR and aar and tested for resistance to Amp by overnight growth on Luria Bertani (LB) agar supplemented with 100 μg/ml Amp. For some patients, we received up to five isolates from the same visit. If those isolates had the same pattern of EAEC genes (aatA, aaiC, aggR, aar, AAF, and SPATEs) we moved forward with only one of the isolates.
The presumptive EAEC strains were grown overnight on LB agar and a single colony was resuspended in 5 mls of LB and grown overnight at 37°C. Genomic DNA (gDNA) was prepared from overnight cultures with the DNeasy Blood and Tissue Kit (Qiagen). The gDNA concentration was quantified with the PicoGreen dsDNA Assay Kit (Thermo Fisher). Sequencing libraries were prepared with 1 ng gDNA input using the Nextera XT standard kit (Illumina FC-121-1024 and FC-121-1001) and the Nextera XT Index Kit v2 (96 indexes for 384 samples). Sequencing libraries were assessed for size distribution and absence of free adapter and adapter dimers using the AATI Fragment Analyzer and library concentration was determined using a quantitative PCR-based assay with the KAPA Library Quantification Kit on the Roche LightCycler 480 Instrument II. The libraries were normalized, pooled and sequenced on a NextSeq 500 (Illumina) using paired-end run conditions for 300 cycles (2x150 with dual index reads). Sequencing run data was demultiplexed using bcl2fastq Conversion Software 2.18 to generate FASTQ files.
Illumina paired-end reads were assembled into larger contigs using the CLC Genomics Workbench platform following the standard procedure. Virulence genes and serotypes were identified from the genome assemblies through the Center for Genomic Epidemiology's Virulence Finder (Joensen et al., 2014), Serotype Finder (Joensen et al., 2015), ResFinder (acquired antibiotic resistance genes, Zankari et al., 2012), and the multi-locus sequence typing (MLST, Larsen et al., 2012) web interfaces, and the Virulence factor database (VFDB) web interface (Liu B. et al., 2019). Presence or absence of virulence genes was further confirmed using CLC Genomics Workbench. The absence of SPATE genes boa (GenBank #AY876285.1), sha (MH899684), tagB and tagC (GenBank # MH899681), and tleA (GenBank # KF494347) was confirmed with Clone Manager 9 software. Putative biofilm-associated genes shf (GenBank # AF134403.1), fis (GenBank # NC_000913.3), yafK (P0AA99.1), and yfaL/ehaC (GenBank # NZ_AFWC01000180.1) were detected with Clone Manager 9 software. Possible adherence genes for AIDA-I (aida, GenBank # JQ044409.1), CS22 (cseA, Genbank # AF145205.1 Pichel et al., 2000, the operons with ecpA (Genbank # FJ210911.1), type 1 fimbriae gene fimA (GenBank # X00981.1), long polar fimbriae operons 1 and 2 (detected by searching for lpf A 1 and 2 from prototype EAEC strain 55989, GenBank # NC_011748.1), and pil [GenBank accession AY686591.1 for pSerB1 (Dudley et al., 2006)], were detected by searching the assembled contigs with BLAST® or Clone Manager 9 software. The sequences from the EAEC from this study are available as part of Bioproject PRJNA576592 in the NCBI BioProject database (https://www.ncbi.nlm.nih.gov/bioproject/).
Fifty of the EAEC genomes sequenced in this study along with the prototypical EAEC reference strain 042 genome (GenBank accession: NC_017626.1, Chaudhuri et al., 2010) were used to construct a whole genome-based phylogenetic tree. The WGS from E15V1A, K6V5, K32V4, K33V5, K29V1, and K36V1were not included in the analyses because we originally were unsure whether to include them because they lacked aggR and aaiC (E15V1A, K29V1, and K36V1—in which we subsequently found a variant aaiC gene), or because they lacked aggR, aap (we later identified an aap-like gene in these strains), and had only one SPATE. The phylogeny was inferred from whole genome alignments (WGA) using Mugsy 1.2.3 (Angiuoli and Salzberg, 2011) and RAxML 4.0 (Stamatakis, 2014) with 100 bootstrap replicates. The tree was visualized in Geneious 2019.1.2 (Kearse et al., 2012) and decorated with strain-associated metadata in Evolview v3 (Zhang et al., 2012; He et al., 2016; Subramanian et al., 2019).
To compute a SNP-based phylogeny for the sampled EAEC, we used a custom-built core genome (cg) SNP discovery pipeline described in more detail in Eppinger et al. (2010, 2011, 2014), Rusconi et al. (2016), and Hau et al. (2018), which is implemented on the open-source web-based bioinformatics platform Galaxy (Goecks et al., 2010). This high-resolution SNP discovery and typing strategy allowed us to put the sampled EAEC strains into a phylogenomic context and determine their individual phylogenetic relationships. We defined the chromosomal core genome as the set of genic and intragenic regions that do not contain phages, repeats, IS elements, plasmid regions, genomic islands, or other mobile genetic elements, which evolve at different rates and are not indicative of evolutionary relationships. These excluded regions were determined for the designated prototypical closed reference EAEC strain 042 (Chaudhuri et al., 2010) as follows: Repeats with NUCmer 3.22 (Delcher et al., 2003), prophages with PHASTER (Arndt et al., 2016, 2017), and IS elements with ISFinder (Siguier et al., 2006), ISEScan 1.7.1 (Xie and Tang, 2017), and ICEberg (Liu M. et al., 2019). The modular read-based SNP discovery pipeline contains the following workflow steps: (i) SNP discovery. Illumina reads of the 50 EAEC query strains were aligned with BWA-MEM (Li and Durbin, 2009) to the designated reference genome EAEC 042. The resulting alignments were processed with Freebayes 1.3.1 (Garrison and Marth, 2012) with the following threshold settings: mapping quality 30, base quality 30, coverage 10, and allelic frequency 0.75. The resulting SNP panel for each of the query genomes was used for further processing; (ii) SNP validation and filtering. We used several SNP curation strategies as described previously (Eppinger et al., 2010, 2011, 2014; Rusconi et al., 2016; Hau et al., 2018). Briefly, cataloged SNPs from each genome were merged into a single SNP panel and SNPs located within excluded regions were removed along with low quality alignments or misalignments, non-uniformly distributed regions and InDels, as previously described (Myers et al., 2009; Morelli et al., 2010; Eppinger et al., 2014). SNPs were further curated by extracting the surrounding nucleotides (40 nt) for each predicted SNP in the reference genome followed by BLASTn search against the query genomes (Altschul et al., 1990); (iii) SNP annotation and chromosomal distribution. Allelic status and chromosomal position of SNPs were recorded and for the biological relevance of the SNPs, polymorphisms were classified into intragenic or intergenic by mapping the SNPs to the 042 reference genome annotation (Manning et al., 2008; Bono, 2009; Chaudhuri et al., 2010; Leopold et al., 2010; Rusconi et al., 2016). In addition, we used a custom-developed genotyper tool to provide a summary of the SNP statistics and report on the number of individual genotypes in the phylogenetic network. (iv) SNP phylogeny. The curated panel of high quality SNPs served as the basis for phylogenetic reconstruction by maximum parsimony with PAUP v4.0a163 (Wilgenbusch and Swofford, 2003) with 100 bootstrap replicates. The majority rule consensus SNP tree was visualized in Geneious (R11.1.5) (Kearse et al., 2012) and decorated in Evolview (Zhang et al., 2012; He et al., 2016; Subramanian et al., 2019). Calculation of the consistency index in Mesquite (Maddison and Maddison, 2019) for each SNP allowed us to identify parsimony informative SNPs and flag homoplastic SNPs, as described previously (Eppinger et al., 2010, 2011, 2014; Rusconi et al., 2016; Hau et al., 2018).
Strains were grown overnight in LB broth with shaking. The following day, 10 μl of overnight culture was added to 1 ml Dulbecco's Modified Eagle Medium (DMEM, ThermoFisher Scientific, Waltham, MA) with 0.45% glucose and L-glutamine and 180 μl of the diluted sample was added in triplicate to a 96-well plate. The samples were incubated for 24 h at 37°C without shaking. After 24 h, the media was carefully aspirated and the biofilms washed once with phosphate-buffered saline (PBS). The biofilms were then fixed with 75% ethanol and allowed to dry completely. The biofilms were then stained with crystal violet, rinsed with water, and allowed to dry. The crystal violet was eluted with 100 μl of 75% ethanol and allowed to incubate for 5 min prior to reading absorbance at a wavelength of 592 nm.
The adherence protocol was based on that of Cravioto et al. (1979). Briefly, HEp-2 cells at a concentration of 2.5 x 105 cells/ml were added to an eight-well chamber slide and incubated overnight in DMEM with 10% fetal bovine serum and penicillin-streptomycin (Lonza) and gentamycin (Quality Biologicals). The same day, cultures of EAEC strains were inoculated into LB and incubated with shaking at 37°C for ~6 h. A 1:100 dilution of the culture was added to DMEM and the culture incubated overnight with shaking at 37°C. The following day, the HEp-2 cells were washed three times with PBS warmed to 37°C, then overlaid with the bacterial inoculum diluted into DMEM with 1% D-mannose for a final multiplicity of infection of 100. The infected HEp-2 cells were incubated for 3 h, then carefully washed three times with PBS, fixed with 0.8% glutaraldehyde/1%formaldehyde in PBS, washed, and stained with 20% Gibco Giemsa Stain solution (Fischer Scientific). Slides were imaged with an Olympus BX60 microscope. The adherence phenotypes were determined by two investigators blinded as to the strain observed according to the adherence descriptions by Nataro et al. (1987).
Mouse studies were conducted in strict accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals. All animal studies were approved by the Institutional Animal Care and Use Committee of the Uniformed Services University of the Health Sciences. Female C57BL/6 mice aged 5–6 weeks from Jackson Labs were used for all mouse experiments. Mice were fasted for ~16 h prior to bacterial inoculation and given drinking water supplemented with 2.5 g/l Amp (Corning). Mice were orally infected with 109 to 1011 CFU in 100 μl by intragastric gavage. The inoculum was prepared from an overnight culture of EAEC grown in LB. The culture was pelleted by centrifugation, and the pellet resuspended in PBS at 1/10 of the original volume. Food was returned ~30 min post-infection (p.i.). Amp treatment continued throughout the course of the experiment. Fresh Amp water was prepared every 48 h. Mice were monitored for weight change for up to 42 days p.i. Weight change was determined as the weight p.i. compared to the weight of the mice the day before infection (d−1). In these studies, the FTT phenotype was assigned to mice that gained less weight compared to PBS mock-infected mice. Weight gain was compared by two way analysis of variance (ANOVA) with Dunnett correction for multiple comparisons. Colonization levels were determined through the course of the experiment, from fecal pellets homogenized in PBS (1:9 weight: volume), then serially diluted and plated onto LB agar plates supplemented with Amp (100 μg/ml).
Formalin-fixed intestine was embedded in paraffin and then sectioned. Tissue sections were then placed on charged glass slides at HistoServ (Germantown, MD). Next, slides were deparaffinized in the Histoclear agent (National Diagnostics, Atlanta, GA) and rehydrated in a graded ethanol series. To increase antibody recognition of the antigen in tissue sections, slides were treated with antigen retrieval buffer [5 × AntigenPlus buffer, pH 10 (EMD Biosciences, San Diego, CA), diluted to 1 ×], heated in a microwave oven for 15 min at 50% power, cooled, and rinsed with water as done previously (Mohawk et al., 2010). Slides were then incubated overnight in blocking buffer (3% bovine serum albumin in PBS). To locate EAEC bound to or near the surface of intestinal tissue, slides with intestinal sections were immunostained with the appropriate anti-O serum (O92 for P73V1 and JM221, O99 for P433V1, Statens Serum Institute, Copenhagen Denmark). The anti-O serum was incubated with the tissue sections at a dilution of 1:50 in PBS for 1 h. The antiserum-exposed tissue sections were then washed, and secondary antibody conjugated to Alexa Fluor 488 was added for 1 h at a dilution of 1:200 in PBS. The slides were rinsed in PBS, 4′,6-diamidino-2-phenylindole (DAPI) Slowfade reagent (ThermoFisher Scientific, Waltham, MA) was added, and a coverslip was applied. Immunofluorescence of stained tissue sections was visualized with an Olympus BX60 microscope with a BX-FLA fluorescent attachment. Digital images of the fluorescent images were obtained with a SPOT RT charge-coupled-device digital camera (Diagnostic Instruments, Inc., Sterling Heights, MI).
We isolated and sequenced gDNA from 56 presumptive EAEC isolates received from field laboratories in the TrEAT TD study. The sequences were assembled into contigs and then analyzed to determine the virulence gene profile and the predicted O and H antigen type. The operons for the known EAEC AAF variants I-V were identified in 35/56 of the strains in the collection (Table 1). The sequence for one strain (E11V1A) contained the genes for both agg3A (entire agg3 operon was present) and agg5A, a phenomenon described previously (Jonsson et al., 2017), although in E11V1A there was only a single agg3B gene. Most of the isolates also had the operons for E. coli common pilus (Ecp), type I fimbriae, long polar fimbriae type 1 or 2, and/or Pil (Table 2). The gene for heat-resistant agglutinin 1, hra1, identified by Bhargava et al. (2009) as an accessory colonization factor, was found in six of the isolates with an AAF operon, and none of the isolates had the gene (aida) for AIDA-I production, an autotransporter reported to confer diffuse adherence in E. coli (Benz and Schmidt, 1989), Table 2. All of the EAEC with an AAF operon had aap, aatA [the entire operon (aatPABCD) was present in all strains that had aatA], and aggR, and, with the exception of the agg4A-positve strains, all had aar (Table 1). The gene that encodes eilA was only found in seven isolates. All of the AAF operon-positive isolates adhered to HEp-2 cells (Table 2).
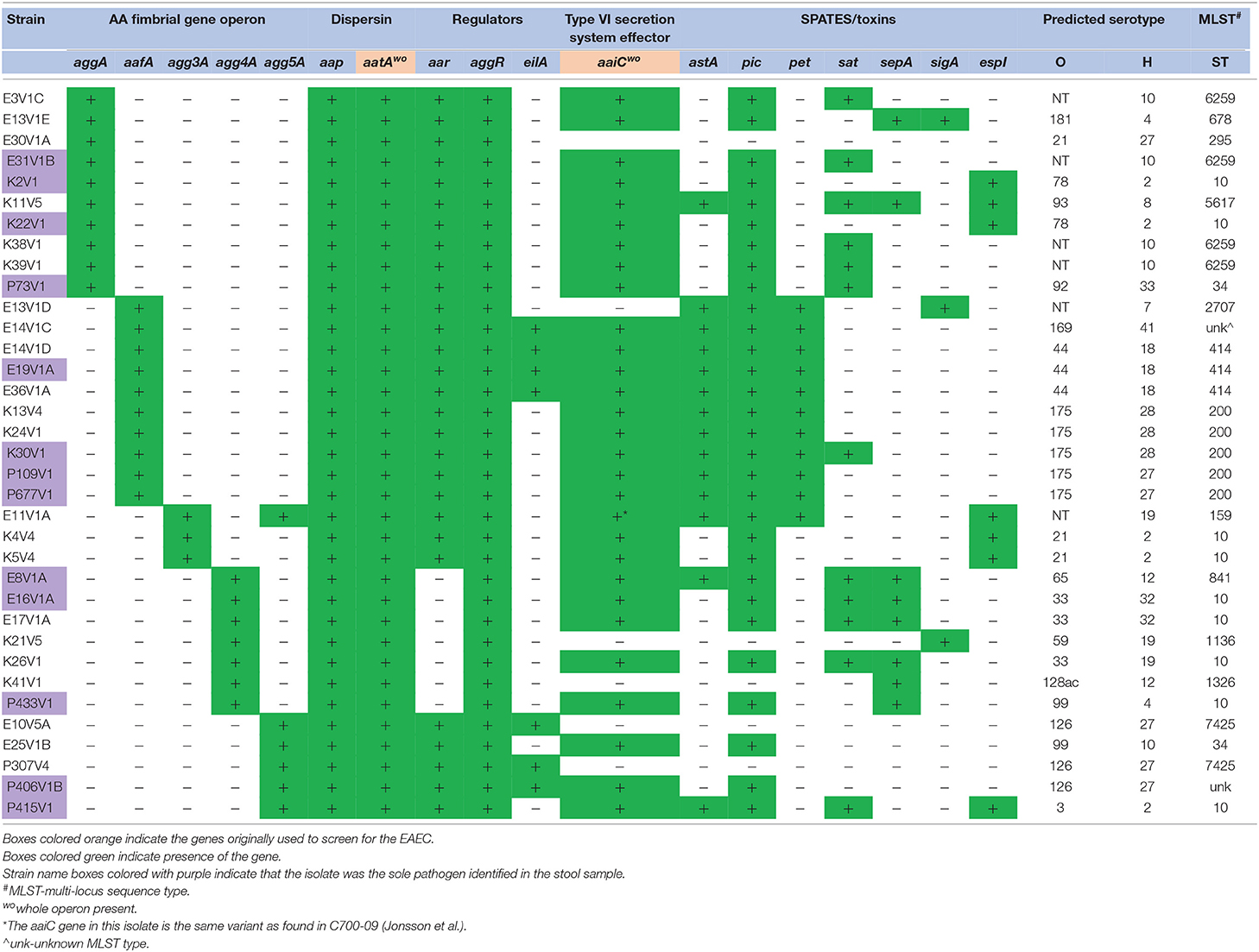
Table 1. Virulence gene profile of EAEC with an aggregative adherence fimbrial gene isolated from DMP.
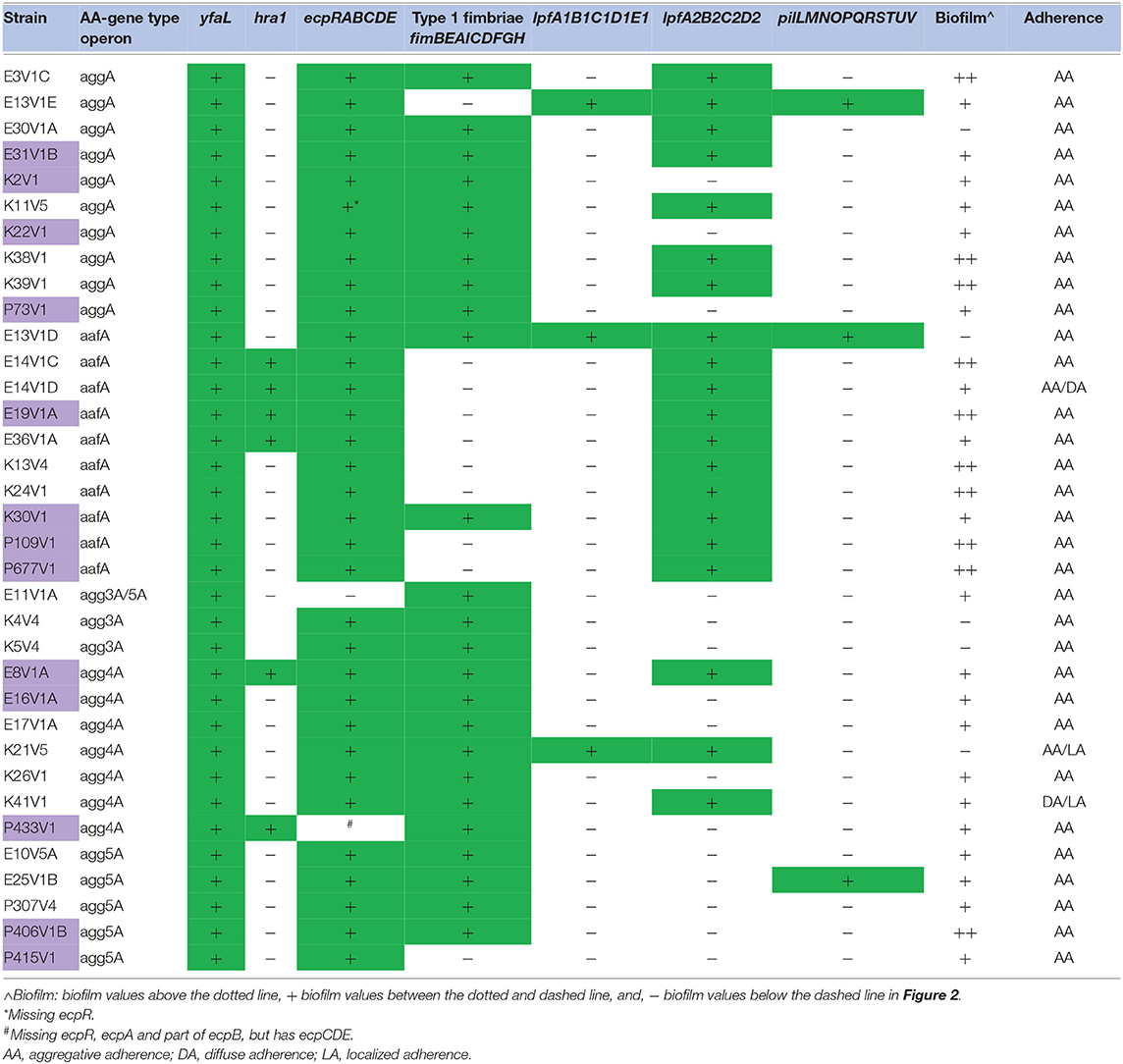
Table 2. Adherence gene or operon presence, and biofilm and adherence phenotype.
All but three of the 35 AAF operon-positive isolates had genes for at least one SPATE (Table 1). Strain E30V1A that lacked any SPATE came from a patient who was also positive for enterotoxigenic E. coli (ETEC), so it is not clear whether the EAEC strain was responsible for the symptoms that patient exhibited. The other two isolates without a SPATE, E10V5A, and P307V4, were isolated during a follow-up visit, and no diarrheal symptoms were reported by the patients at that time.
An AAF operon was not identified for 21/56 (37.5%) presumptive EAEC isolates (Table 3). Similar rates for a lack of an AAF gene in EAEC were previously reported (Boisen et al., 2012; Jonsson et al., 2015). Nine of the 21 strains without a known EAEC AAF gene had a gene almost identical (97–99% homologous) to the gene (cseA) for colonization factor CS22 characterized in enterotoxigenic E. coli (ETEC) strains (Pichel et al., 2000). The presence of an operon for CS22 was recently identified in other EAEC (Boisen personal communication), and is also present in the cseA+ strains identified in this study (Supplemental Figure 1). All of the cseA-positive strains had aggR and at least one SPATE (Table 3), but lacked the genes for ETEC heat-stabile or heat labile toxin genes. However, we found that the isolates that have the CS22-like operon did not adhere to HEp-2 cells in our hands or showed minimal (a few bacteria could be found adherent to the HEp-2 cells) adherence (Table 4). ETEC with CS22 have been shown to adhere to Caco-2 cells (Pichel et al., 2000), but we did not test our strains on Caco-2 cells.
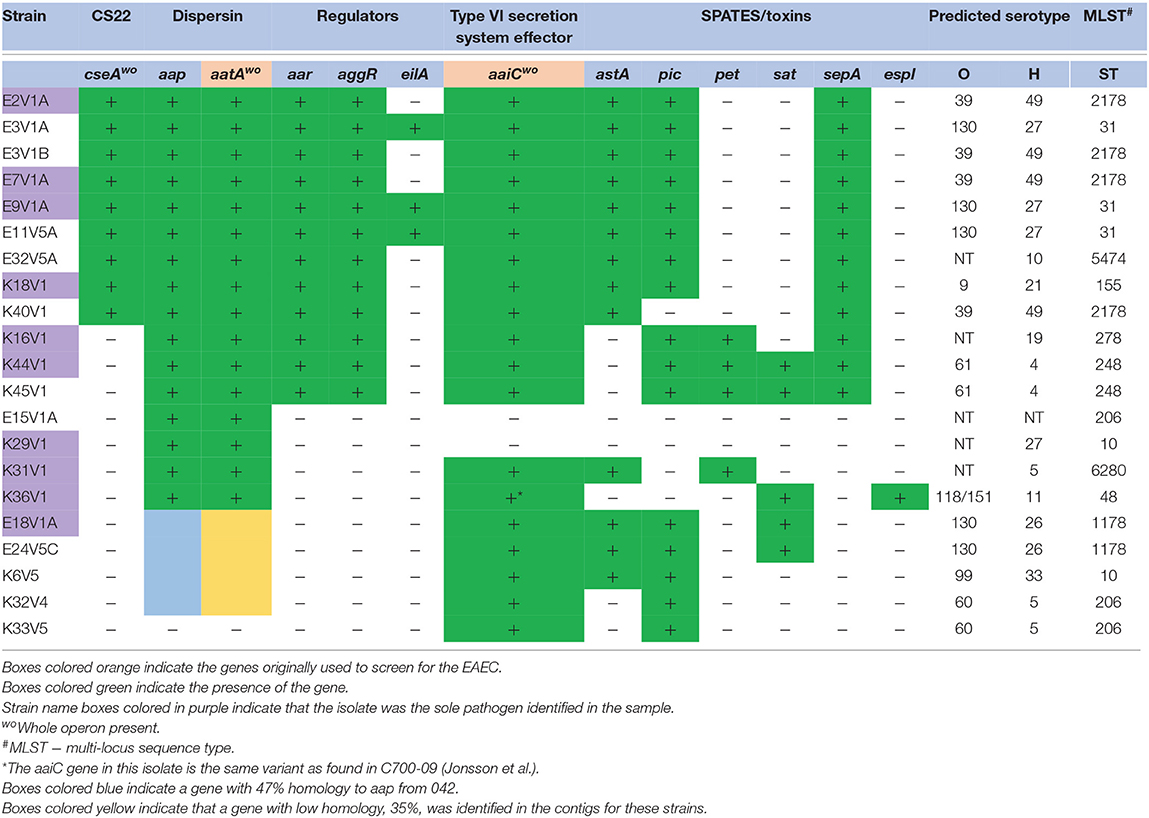
Table 3. Virulence gene profile of E. coli from DMP that lacked an AA fimbrial gene.

Table 4. Adherence genes or operons andbiofilm and adherence phenotype for strains without an AA-fimbrial gene operon.
There were 12 strains without an AAF gene or the CS22-like operon (Tables 3, 4). Of those isolates, K16V1, K44V1, and K45V1 had other adhesin gene markers, as well as aap, attA, aar, aggR, and three or more SPATEs, genotypes which marked them as EAEC, although they did not encode an AAF gene. We were unable to locate adhesin genes on the same contigs as genes that are typically found on the pAA plasmid (aap, attA, aar, aggR, pet, and sepA) in these latter strains. Of the final nine isolates listed in Table 3 without a known AAF gene or the CS22 operon, all lacked aggR and aar as well, but seven of those did have at least one SPATE. Eight of these nine isolates had the ecp operon that encodes the E. coli common pilus, an operon shown to be in other EAEC (Avelino et al., 2010). However, many of the strains that had the ecp operon did not adhere well to HEp-2 cells, which indicates that the presence of the ecp operon does not correlate with adherence. In contrast, two of the AAF gene-negative ecp operon-positive strains, E18V1A, and K6V5 did adhere well to HEp-2 cells (Table 4). The isolate (K31V1) that lacked an EAEC AAF gene, the CS22-like operon, lpf , or ecp, had the pil operon. In this collection, we found the pil operon in nine isolates without AAF operons and three of the 35 strains that had an EAEC AAF operon. However, the pil+ strains that were AAF gene-negative did not adhere well to HEp-2 cells in this study. Finally, none of the 21 isolates without an AAF operon had the gene (aida) for AIDA-I (associated with diffuse adherence).
All but 5/21 of the isolates without an AAF gene had the structural gene for dispersin, aap, and the dispersin transport system gene used for the original stool screening, aatA (Table 3), and in fact encode the entire operon for dispersin transport (aatPABCD). The five strains that lacked aap also lacked aggR and other genes that are markers for the pAA plasmid (Table 3). We suspected that one or more of these five strains may have lost the large plasmid during isolation, or carry a different plasmid, so we searched for other possible markers for the EAEC pAA. From that search, we found that 4/5 strains had a gene with 47% amino acid identity to aap (Supplemental Figure 2), and genes homologous to aatPABCD, with a lower overall identity that ranged from 30 to 47% depending on which gene in the operon was compared (not shown). The dispersin-like and dispersin-transport-like gene(s) were found in only one of the EAEC with an AAF gene, P406V1B. P406V1B also has the genes for the traditional dispersin and dispersin transport.
We next considered only those 23 EAEC isolates that came from visit 1 (sick visit) and which were the only identified pathogen from the stool (Table S1). The 23 isolates were 96% positive for aap, aatA, and aaiC and all had yfaL. The next most common genes were pic (87%), aggR (83%), and aar (70%). We also included the maximum number of stools in a 24 h period from the patients infected with the isolates listed in Table S1. The range for the number of stools from these isolates was 3–10, and the number from the isolates with cseA (4–7) was similar. In fact, no differences in the clinical picture from the patients with cseA isolates were noted overall.
To search for associations between EAEC genes present in any visit 1 (sick visit) isolate as compared to convalescent visits 4 and 5, we calculated the odds ratios (OR, Table 3). We found that the presence of app was associated with visit 1 (sick visit) as compared to visits 4 and 5 (follow-up visits), with an OR of 18.7 (P = 0.008), though the 95% confidence intervals were wide. None of the other EAEC genes correlated with visit 1 as compared to visits 4 and 5. We did a similar analysis for the EAEC that were identified as the only pathogen in the diarrheal stool compared to stool with EAEC and an additional pathogen(s), but found no correlations (Table 5).
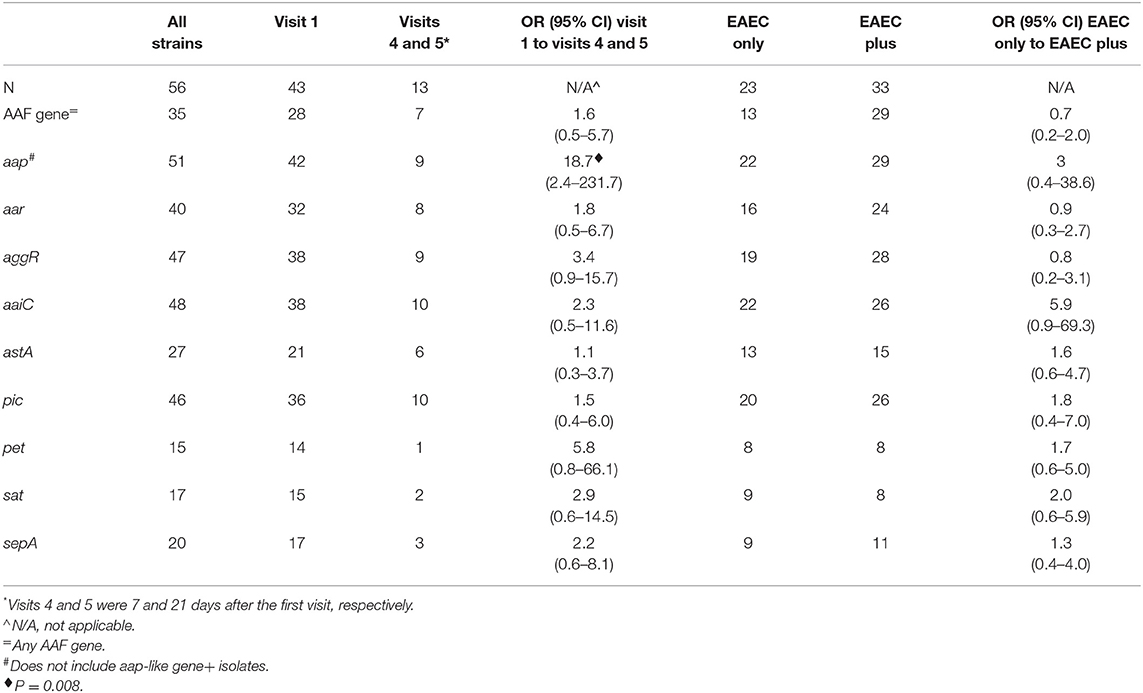
Table 5. Odds ratios for the presence of EAEC genes in isolates from visit 1 compared to visits 4 and 5 and in isolates identified as the only pathogen compared to those which were not the only pathogen.
Over the entire collection of 56 isolates, there were a variety of O and H genotypes identified (Tables 1, 3). There were 21 predicted O antigen types with O130 and O175 present 5 times. The O antigen could not be typed in 11 strains. For the H antigen, the genes for H27, H6, and H2 were the most common. Some O:H combinations were more common than others: ONT:H10 (five strains), O39:H49 (four strains), O44:H18 (three strains), O126:H27 (three strains), O130:H27 (three strains), and O175:H28 (three strains). Notably, the serotypes for EAEC prototype strains JM221 (O92:H33) and 042 (O44:H18) were found in our collection. Finally, E14V1C was O169:H41, a serotype that has been linked to ETEC outbreaks in Asia and the U.S. (Beatty et al., 2004; Harada et al., 2013). Isolate E14V1C does have the heat stable enterotoxin (ST) gene (the only strain in the collection with that toxin gene) and may be a hybrid EAEC/ETEC strain. As expected for EAEC the strains had a variety of antibiotic resistance genes (Table S2).
We found that three patients appeared to be infected with two (E13V1D and E13V1E; E14V1C and E14V1D) or three (E3V1 A, B, and C) EAEC which differed by virulence gene make-up and/or serotype genes. This latter finding indicates that individuals may be infected with multiple EAEC at the same time, and demonstrates the importance of having multiple individual isolates from patients.
A phylogenetic tree was established by aligning the whole genomes of 50 EAEC isolated from the DMP along with the genome of the prototype EAEC strain 042 (Figure 1). The WGA tree topology shows that the strains are separated into different genetic clusters and we observed correlations to AAF gene type, the predicted O and/or H antigen type, and somewhat by SPATE genes sepA, sigA, and the regulator eilA. The overall topology of the WGA phylogeny is largely mirrored by the SNP inferred phylogeny (Supplemental Figure 3). The SNP discovery yielded a total of 155,432 core genome SNPs (Table S3), of which 141,761 were parsimony informative. The latter is indicative of the diversity in the sampled EAEC strains, and is also reflected in the many branch points and individual clusters. However, in some cases EAEC from different patients in the same geographical area were found to be highly related and separated by fewer than 80 core genome SNPs from each other. Examples of such isolates include K38V1 and K39V1 (28 SNPs); E16V1A and E17V1A (55 SNPs); and E14V1D and E37V1A (6 SNPs) which were collected within the same day or week (see Supplemental Figure 3, black boxes). In contrast, other sets of genotypically highly similar isolates came from patients who reported illness 5 months [E3V1A and E11V5A (17 SNPs); K13V4 and K24V1 (34 SNPs)] or 1 year [E3V1C and E31V1B (79 SNPs)] apart (Supplemental Figure 3, green boxes). Further, there were two strains with just 46 SNPs from each other that were isolated from different DMP more than 1 year apart on different continents (E10V5A and P307V4, Supplemental Figure 3, red box). We also found examples of isolates that were more distantly related isolated from patients on the same day or 1 day apart [E7V1A compared to E8V1A (18,832 SNPs); E37V1A compared to the E13 strains (more than 55,600 SNPs); K40V1 compared to K38V1 and K39V1 (more than 19,100 SNPs)], findings that indicate that DMP may be exposed to multiple types of EAEC in the same geographic location.
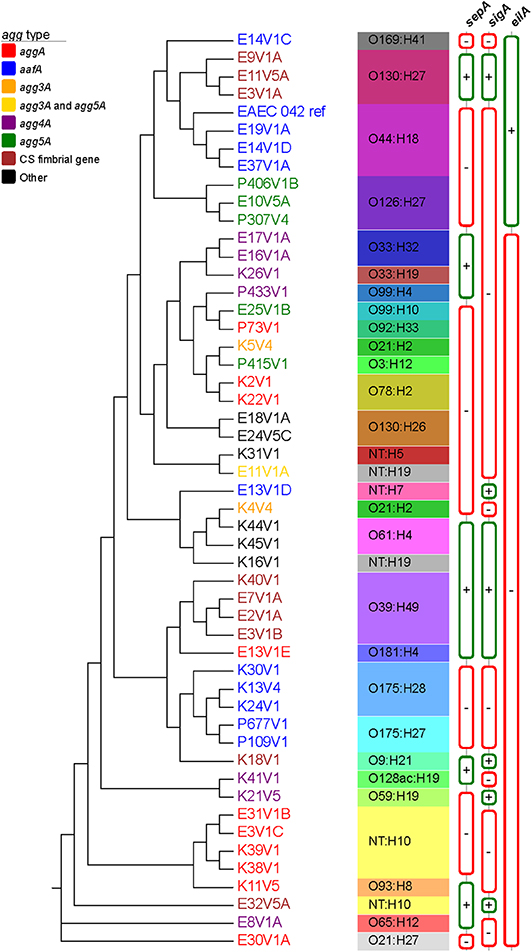
Figure 1. Whole genome phylogeny of sampled EAEC strains. The complete genomes of a total of 51 EAEC strains, which included the prototypical EAEC reference genome 042 (Chaudhuri et al., 2010), were aligned with Mugsy (Angiuoli and Salzberg, 2011). The phylogenetic tree was inferred using RAxML (Stamatakis, 2014) with 100 bootstrap replicates and decorated with the metadata in Evolview (Zhang et al., 2012; He et al., 2016; Subramanian et al., 2019). AAF type is depicted with different leaf colors, as shown in the legend. Serotype as well as sepA, sigA, and eilA status were also included in the tree. All nodes presented bootstrap values of >90. The tree topology partitions the isolates into distinct phylogenetic clusters in accordance with strain associated metadata.
In vitro biofilm formation after 24 h was assessed for all EAEC strains (Figure 2, Tables 2, 4). In addition to the EAEC DMP strain collection, we used K12 strain MG1655 as a negative control, and prototype EAEC JM221 as a positive control. The biofilm-staining data in Figure 2 were grouped by fimbrial gene type and from the highest to lowest biofilm former to make the data easier to read. Within each fimbrial genotype we found variation in biofilm formation capacity, and a few strains did not form a biofilm by this assay (Figure 2). We found three strains (E18V1A, E24V5C, and K6V5) that lacked an AAF gene/operon were capable of moderate biofilm formation. The remainder of the strains without an AAF gene/operon were unable to form a biofilm (data not shown). All of the strains in this study had yfaL/ehaC, which if overexpressed in E. coli K-12 leads to enhanced biofilm formation (Roux et al., 2005), and fis, yafK, and shf, which if mutated, reduce biofilm formation (Sheikh et al., 2001; Fujiyama et al., 2008). However, all of the strains in this study were missing the start codon (ATG) of shf , so if the gene is expressed, the protein is likely truncated.
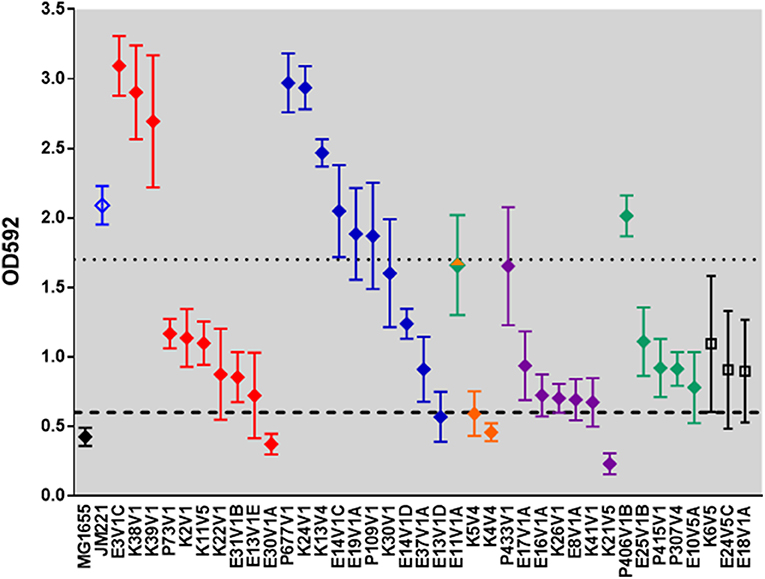
Figure 2. Biofilm-formation capacity of clinical isolates. The data represent the mean OD592 from at least three independent biofilm assays. The black diamond represents E. coli K12 strain MG1655. EAEC prototype strain JM221 is shown in blue. The filled diamonds are colored according to AAF operon type: aggA (red), aafA (blue), agg3A (orange), agg4A (purple), agg5a (green), and no AAF gene (black). Strain E11V1 is agg3a+ & agg5a+ and is shown with an orange and green diamond. Strains K6V5, E24V5C, and E18V1A did not have an AAF gene, and are indicated by open black squares. The dashed line is one standard deviation above the mean value for MG1655, and the dotted line is one standard deviation below the mean value for JM221. We classified high biofilm formers as those with values above the dotted line, and non-biofilm formers as those with values below the dashed line.
We used an Amp-treated mouse model to assess the pathogenesis of a subset of the EAEC collection. Because the Amp-treated mouse model was originally developed to characterize the O104:H4 2011 Shiga toxin-producing E. coli outbreak strain C227-11 (Zangari et al., 2013), and none of the EAEC strains in this collection had genes for Shiga toxin, we first tested the prototype EAEC strain JM221 in the model. We observed that mice infected with JM221 gained less weight over the course of the experiment compared to mock-infected controls, data not shown. We next tested a subset of EAEC from our DMP collection in the Amp-treated model. We began with Amp-resistant isolates from the Peru collection because those were the isolates we received first. Amp-treated mice infected with P73V1, P415V1, or P677V1 gained weight similar to that observed in the mock-infected controls (Figure 3A). However, mice infected with strain P433V1 (agg4A+ aar–) displayed significant FTT when compared to mock-infected controls (Figure 3A). We then tested P433V1 in a larger group of mice and again observed an FTT phenotype (Figure 3B). We also tested other agg4A+ and aar– strains in the mice. In a separate study, we found that strain K26V1, but not E17V1A caused a mild FTT phenotype (Figure 3C).

Figure 3. DMP strain P433V1 caused FTT in an Amp-treated adult-mouse model. (A) Mean weight change of mice infected with EAEC or mock-infected with PBS. Weights were compared with two-way ANOVA. Mice infected with P433V1 (A) showed differences in weight on day 10 and days 14–28 (P = 0.033 for days 10, 14, 28; 0.002 for days 17, 19, 21, and 26; and 0.0002 for day 24). There was no significant difference in weight gain between the other DMP strains (P73V1, P415V1, or P677V1) and the mock-infected mice. (B) Infection with strain P433V1 produced a reproducible FTT phenotype when compared to mock-infected mice. (P = 0.002 for day 20; 0.0001 for days 8, 11, and 18; <0.0001 for days 13, 16, 23, and 27 (n = 5 for mock-infected mice and n = 10 for P433V1). (C) Mean weight of mice (n = 5) infected with K26V1, E17V1A, or mock-infected with PBS. Mice infected with K26V1 showed differences in weight on days 20 and 27 (P = 0.015 for day 20 and 0.0001 for day 27). There was no significant difference in weight gain between E17V1A and mock-infected mice.
While we were unable to identify an in vitro genotype unique for the isolates that caused FTT in the Amp-treated mice, we wanted to determine if the strains displayed differences in the capacity to form a biofilm in vivo. Intestinal sections from mice infected with JM221, P433V1, or P73V1 showed that there were high levels of bacteria in the sections from the mice that caused FTT (JM221 and P433V1), Figure 4, but that the sections from mice infected with P73V1 (did not cause FTT) had very few bacteria present. The colonization levels in the mice as determined by shedding into fecal samples was consistent throughout the course of the experiment among all strains at about 109 CFU/g feces (not shown).
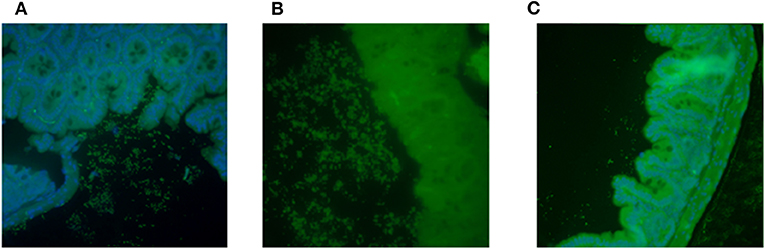
Figure 4. Capacity to form biofilm in vivo correlated with FTT phenotype. Intestines from EAEC-infected mice were stained with fluorescent antibody against the appropriate O-antigen. Mice infected with JM221 (A) or P433V1 (B) showed high levels of bacteria near the intestinal epithelium. Very few bacteria were observed in mice infected with P73V1 (C). Sections were stained from three mice and representative images are shown.
We set out to identify an EAEC genotype associated with TD in DMP. Although we did not identify a single such genotype, we did find a link between the presence of aap in the EAEC isolated at the acute clinic visit as compared to follow-up visits. Similarly, a study in Danish children found a link between aap and acute (as opposed to persistent) diarrhea in children (Hebbelstrup Jensen et al., 2017). The reduced number of EAEC with aap isolated from follow-up visits could indicate that strains without aap are less pathogenic than those with aap, and/or that the patients had mounted an immune response to dispersin. Therefore, it may be that dispersin would be a candidate vaccine antigen for EAEC. However, because we only had 12 isolates from follow-up visits in our study, the finding of strains that lack aap in patients without clinical disease needs to be confirmed. Finally, we found that some of the agg4A positive isolates caused FTT in an Amp-treated mouse model.
Our collection of EAEC isolates from DMP demonstrated genetic heterogeneity for the genes for AAF, other adhesins, and the SPATEs, with the exception of pic and gene yfaL which were found in >80% or all of the isolates, respectively. Most strains were also positive for aggR, aar, aatPABCD, aap, and aaiA-Y. One notable exception was that the agg4A strains lacked aar, a finding observed for other agg4A+ strains as well (Boisen, personal communication). For the SPATEs, the clearest associations were that the aafA strains had pet (a finding also noted by Lima et al., 2013), and all of the isolates with the CS22-like operon and most of the agg4A strains were sepA positive. The WGA and SNP analyses showed the genetic diversity among the EAEC, with many branch points and thousands of SNPs among the strains (see Table S1). However, the SNP analysis also demonstrated that some of the isolates were highly related with as few as 6 SNPs from each other in the core genome, suggesting a common source. These highly related strains came from patients who presented either within the same week or month, but a few were isolated more than a year apart. The finding of such genetically closely-linked isolates from patients who presented a year apart indicates that at least some of the strains are stable in that geographic region, perhaps re-circulating among the population. Furthermore, we ascertained that while most patients had only clonal isolates recovered, some patients were infected with multiple EAEC. These data collectively support the conclusion that many different EAEC are present in the same region. Another set of strains were just 46 SNPs apart in the core genome, but were isolated on different continents, a finding that suggests global distribution of EAEC.
We were somewhat surprised by the number of isolates that lacked an AAF gene that were associated with clinical disease and were the sole identified pathogen (Table 2, purple boxes and Table S1). This latter finding suggests that AAF-negative EAEC strains are associated with TD in this population of patients. It is also possible that some of the AAF-negative isolates did have a pAA plasmid at some point, but that it was lost during cultivation. However, the argument against that possibility is that many of these strains also had genes that are associated with pAA, such as aggR, aap, pet, and sepA. Therefore, it may be that the operon for CS22 replaced the AAF gene operon in these strains. Taken together these data suggest that the strains with the CS22-like fimbrial operon are EAEC that have acquired ETEC adherence genes. Finally, almost all of the isolates without an AAF gene had aaiC, a gene associated with EAEC.
We identified two strains that may have a novel adhesin mechanism as they lacked AAF genes, hra1, and lpf , and pil operons but adhered to HEp-2 cells. Although both strains did encode the ecp operon, and yfaL, the presence of ecp and yfaL did not correlate with adherence in our strains overall. These latter two isolates and two additional strains had a gene with high identity to aap (dispersin) and the operon for dispersin transport. The significance of the dispersin-like genes is not clear, but they could potentially contribute to adherence and/or biofilm formation in these strains.
Finally, we used a mouse model of infection established in our laboratory (Zangari et al., 2013; Boisen et al., 2019) to assess if a subset of the clinical isolates would be associated with morbidity in the animals. We found that Amp-treated mice infected with P433V1 or K26V1 showed FTT compared to mock-infected animals. Furthermore, P433V1 formed a strong biofilm in vitro and in vivo. Both P433V1 and K26V1 have agg4A and sepA but lack aar. A previous study showed an association between sepA and pediatric diarrhea cases compared to control patients (Boisen et al., 2012), and an agg4A+ Shiga toxin-producing E. coli (STEC)/EAEC hybrid strain that caused hemolytic uremic syndrome was described recently (Carbonari et al., 2019). However, the individual contributions of SepA and Agg4A in the disease process remain to be elucidated. In summary, we found that DMP with TD had EAEC that appeared to be similar to EAEC isolated from populations in other settings.
The datasets generated for this study can be found in the NCBI Bioproject, accession number PRJNA576592.
The animal study was reviewed and approved by Uniformed Services University Institutional Animal Care and Use Committee.
CP, JD, YS, and AM-C conceived, planned, and carried the experiments with the support of all other authors. CP and AM-C wrote the manuscript with assistance from AA-G and ME. CD and GS provided experimental support and analysis, specifically with the sequencing. MR, DT, RJ, PC, and NB provided critical feedback and helped to shape the research and analysis.
This work was supported by an intramural program project grant, HU0001-14-1-0061, to MR, DT, and AM-C. The contributions of AA-G and ME were supported by the US Department of Homeland Security [2014-ST-062-000058] and NIAID [SC2AI120941] to ME.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The opinions or assertions contained herein are the private ones of the authors and are not to be construed as official or reflecting the views of the Department of Defense, the Uniformed Services University of the Health Sciences, or the National Institutes of Health. We thank Alison D. O'Brien for her excellent mentorship and review of the manuscript. We thank Anthony Soc for his technical assistance with adherence and biofilm assays.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2020.00200/full#supplementary-material
Al-Hasani, K., Henderson, I. R., Sakellaris, H., Rajakumar, K., Grant, T., Nataro, J. P., et al. (2000). The sigA gene which is borne on the she pathogenicity island of Shigella flexneri 2a encodes an exported cytopathic protease involved in intestinal fluid accumulation. Infect. Immun. 68, 2457–2463. doi: 10.1128/IAI.68.5.2457-2463.2000
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Angiuoli, S. V., and Salzberg, S. L. (2011). Mugsy: fast multiple alignment of closely related whole genomes. Bioinformatics 27, 334–342. doi: 10.1093/bioinformatics/btq665
Arndt, D., Grant, J. R., Marcu, A., Sajed, T., Pon, A., Liang, Y., et al. (2016). PHASTER: a better, faster version of the PHAST phage search tool. Nucleic Acids Res. 44, W16–21. doi: 10.1093/nar/gkw387
Arndt, D., Marcu, A., Liang, Y., and Wishart, D. S. (2017). PHAST, PHASTER and PHASTEST: tools for finding prophage in bacterial genomes. Brief. Bioinform. 20, 1560–1567. doi: 10.1093/bib/bbx121
Avelino, F., Saldana, Z., Islam, S., Monteiro-Neto, V., Dall'agnol, M., Eslava, C. A., et al. (2010). The majority of enteroaggregative Escherichia coli strains produce the E. coli common pilus when adhering to cultured epithelial cells. Int. J. Med. Microbiol. 300, 440–448. doi: 10.1016/j.ijmm.2010.02.002
Beatty, M. E., Bopp, C. A., Wells, J. G., Greene, K. D., Puhr, N. D., and Mintz, E. D. (2004). Enterotoxin-producing Escherichia coli O169:H41, United States. Emerg. Infect. Dis. 10, 518–521. doi: 10.3201/eid1003.030268
Benjelloun-Touimi, Z., Sansonetti, P. J., and Parsot, C. (1995). SepA, the major extracellular protein of Shigella flexneri: autonomous secretion and involvement in tissue invasion. Mol. Microbiol. 17, 123–135. doi: 10.1111/j.1365-2958.1995.mmi_17010123.x
Benz, I., and Schmidt, M. A. (1989). Cloning and expression of an adhesin (AIDA-I) involved in diffuse adherence of enteropathogenic Escherichia coli. Infect. Immun. 57, 1506–1511. doi: 10.1128/IAI.57.5.1506-1511.1989
Bhargava, S., Johnson, B. B., Hwang, J., Harris, T. A., George, A. S., Muir, A., et al. (2009). Heat-resistant agglutinin 1 is an accessory enteroaggregative Escherichia coli colonization factor. J. Bacteriol. 191, 4934–4942. doi: 10.1128/JB.01831-08
Boisen, N., Melton-Celsa, A. R., Hansen, A. M., Zangari, T., Smith, M. A., Russo, L. M., et al. (2019). The role of the AggR regulon in the virulence of the Shiga toxin-producing enteroaggregative Escherichia coli epidemic O104:H4 strain in mice. Front. Microbiol. 10:1824. doi: 10.3389/fmicb.2019.01824
Boisen, N., Scheutz, F., Rasko, D. A., Redman, J. C., Persson, S., Simon, J., et al. (2012). Genomic characterization of enteroaggregative Escherichia coli from children in Mali. J. Infect. Dis. 205, 431–444. doi: 10.1093/infdis/jir757
Bono, J. L. (2009). Genotyping Escherichia coli O157:H7 for its ability to cause disease in humans. Curr. Protoc. Microbiol. Chapter 5:Unit 5A.3. doi: 10.1002/9780471729259.mc05a03s14
Canizalez-Roman, A., and Navarro-Garcia, F. (2003). Fodrin CaM-binding domain cleavage by pet from enteroaggregative Escherichia coli leads to actin cytoskeletal disruption. Mol. Microbiol. 48, 947–958. doi: 10.1046/j.1365-2958.2003.03492.x
Carbonari, C. C., Ricciardi, M., Calvo, A. R., Montes, A., Deza, N. L., Conde Valentino, M. A., et al. (2019). An Stx-EAEC O59:NM[H19] strain isolated from a hemolytic uremic syndrome case in Argentina. Rev Argent Microbiol. 52, 31–35. doi: 10.1016/j.ram.2019.03.006
Chaudhuri, R. R., Sebaihia, M., Hobman, J. L., Webber, M. A., Leyton, D. L., Goldberg, M. D., et al. (2010). Complete genome sequence and comparative metabolic profiling of the prototypical enteroaggregative Escherichia coli strain 042. PLoS ONE 5:e8801. doi: 10.1371/journal.pone.0008801
Cravioto, A., Gross, R. J., Scotland, S., and Rowe, B. (1979). An adhesive factor found in strains of Escherichia coli belonging to the traditional infantile enteropathogenic serotypes. Curr. Microbiol. 3, 95–99. doi: 10.1007/BF02602439
Delcher, A. L., Salzberg, S. L., and Phillippy, A. M. (2003). Using MUMmer to identify similar regions in large sequence sets. Curr. Protoc. Bioinformatics Chapter 10:Unit 10.3. doi: 10.1002/0471250953.bi1003s00
Dudley, E. G., Abe, C., Ghigo, J. M., Latour-Lambert, P., Hormazabal, J. C., and Nataro, J. P. (2006). An IncI1 plasmid contributes to the adherence of the atypical enteroaggregative Escherichia coli strain C1096 to cultured cells and abiotic surfaces. Infect. Immun. 74, 2102–2114. doi: 10.1128/IAI.74.4.2102-2114.2006
Eppinger, M., Mammel, M. K., Leclerc, J. E., Ravel, J., and Cebula, T. A. (2011). Genomic anatomy of Escherichia coli O157:H7 outbreaks. Proc. Natl. Acad. Sci. U.S.A. 108, 20142–20147. doi: 10.1073/pnas.1107176108
Eppinger, M., Pearson, T., Koenig, S. S., Pearson, O., Hicks, N., Agrawal, S., et al. (2014). Genomic epidemiology of the haitian cholera outbreak: a single introduction followed by rapid, extensive, and continued spread characterized the onset of the epidemic. MBio 5:e01721. doi: 10.1128/mBio.01721-14
Eppinger, M., Worsham, P. L., Nikolich, M. P., Riley, D. R., Sebastian, Y., Mou, S., et al. (2010). Genome sequence of the deep-rooted Yersinia pestis strain angola reveals new insights into the evolution and pangenome of the plague bacterium. J. Bacteriol. 192, 1685–1699. doi: 10.1128/JB.01518-09
Fujiyama, R., Nishi, J., Imuta, N., Tokuda, K., Manago, K., and Kawano, Y. (2008). The Shf gene of a Shigella flexneri homologue on the virulent plasmid pAA2 of enteroaggregative Escherichia coli 042 is required for firm biofilm formation. Curr. Microbiol. 56, 474–480. doi: 10.1007/s00284-008-9115-y
Garcia, B. G., Castro, F. S., Vieira, M. A. M., Girao, D. M., Uenishi, L. T., et al. (2019). Distribution of the pilS gene in Escherichia coli pathovars, its transfer ability and influence in the typical enteropathogenic E. coli adherence phenotype. Int. J. Med. Microbiol. 309, 66–72. doi: 10.1016/j.ijmm.2018.12.001
Garrison, E., and Marth, G. (2012). Haplotype-Based Variant Detection From Short-Read Sequencing. ArXiv e-prints [Online], 1207. Available online at: http://adsabs.harvard.edu/abs/2012arXiv1207.3907G (accessed July 1, 2012).
Goecks, J., Nekrutenko, A., Taylor, J., and Galaxy, T. (2010). Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 11:R86. doi: 10.1186/gb-2010-11-8-r86
Guignot, J., Chaplais, C., Coconnier-Polter, M. H., and Servin, A. L. (2007). The secreted autotransporter toxin, sat, functions as a virulence factor in Afa/Dr diffusely adhering Escherichia coli by promoting lesions in tight junction of polarized epithelial cells. Cell. Microbiol. 9, 204–221. doi: 10.1111/j.1462-5822.2006.00782.x
Harada, T., Itoh, K., Yamaguchi, Y., Hirai, Y., Kanki, M., Kawatsu, K., et al. (2013). A foodborne outbreak of gastrointestinal illness caused by enterotoxigenic Escherichia coli serotype O169:H41 in Osaka, Japan. Jpn. J. Infect. Dis. 66, 530–533. doi: 10.7883/yoken.66.530
Harrington, S. M., Sheikh, J., Henderson, I. R., Ruiz-Perez, F., Cohen, P. S., and Nataro, J. P. (2009). The pic protease of enteroaggregative Escherichia coli promotes intestinal colonization and growth in the presence of mucin. Infect. Immun. 77, 2465–2473. doi: 10.1128/IAI.01494-08
Hau, S. J., Allue-Guardia, A., Rusconi, B., Haan, J. S., Davies, P. R., Frana, T. S., et al. (2018). Single nucleotide polymorphism analysis indicates genetic distinction and reduced diversity of swine-associated methicillin resistant Staphylococcus aureus (MRSA) ST5 isolates compared to clinical MRSA ST5 isolates. Front. Microbiol. 9:2078. doi: 10.3389/fmicb.2018.02078
Havt, A., Lima, I. F., Medeiros, P. H., Clementino, M. A., Santos, A. K., Amaral, M. S., et al. (2017). Prevalence and virulence gene profiling of enteroaggregative Escherichia coli in malnourished and nourished Brazilian children. Diagn. Microbiol. Infect. Dis. 89, 98–105. doi: 10.1016/j.diagmicrobio.2017.06.024
He, Z., Zhang, H., Gao, S., Lercher, M. J., Chen, W. H., and Hu, S. (2016). Evolview v2: an online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 44, W236–241. doi: 10.1093/nar/gkw370
Hebbelstrup Jensen, B., Poulsen, A., Hebbelstrup Rye Rasmussen, S., Struve, C., Engberg, J. H., Friis-Moller, A., et al. (2017). Genetic virulence profile of enteroaggregative Escherichia coli strains isolated from danish children with either acute or persistent diarrhea. Front. Cell. Infect. Microbiol. 7:230. doi: 10.3389/fcimb.2017.00230
Joensen, K. G., Scheutz, F., Lund, O., Hasman, H., Kaas, R. S., Nielsen, E. M., et al. (2014). Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 52, 1501–1510. doi: 10.1128/JCM.03617-13
Joensen, K. G., Tetzschner, A. M., Iguchi, A., Aarestrup, F. M., and Scheutz, F. (2015). Rapid and easy in silico serotyping of Escherichia coli isolates by use of whole-genome sequencing data. J. Clin. Microbiol. 53, 2410–2426. doi: 10.1128/JCM.00008-15
Jonsson, R., Struve, C., Boisen, N., Mateiu, R. V., Santiago, A. E., Jenssen, H., et al. (2015). Novel aggregative adherence fimbria variant of enteroaggregative Escherichia coli. Infect. Immun. 83, 1396–1405. doi: 10.1128/IAI.02820-14
Jonsson, R., Struve, C., Boll, E. J., Boisen, N., Joensen, K. G., Sorensen, C. A., et al. (2017). A novel pAA virulence plasmid encoding toxins and two distinct variants of the fimbriae of enteroaggregative Escherichia coli. Front. Microbiol. 8:263. doi: 10.3389/fmicb.2017.00263
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., et al. (2012). Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649. doi: 10.1093/bioinformatics/bts199
Larsen, M. V., Cosentino, S., Rasmussen, S., Friis, C., Hasman, H., Marvig, R. L., et al. (2012). Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 50, 1355–1361. doi: 10.1128/JCM.06094-11
Leopold, S. R., Shaikh, N., and Tarr, P. I. (2010). Further evidence of constrained radiation in the evolution of pathogenic Escherichia coli O157:H7. Infect. Genet. Evol. 10, 1282–1285. doi: 10.1016/j.meegid.2010.07.021
Li, H., and Durbin, R. (2009). Fast and accurate short read alignment with burrows-wheeler transform. Bioinformatics 25, 1754–1760. doi: 10.1093/bioinformatics/btp324
Lima, I. F., Boisen, N., da Quetz, J. S, Havt, A., De Carvalho, E. B., Soares, A. M., et al. (2013). Prevalence of enteroaggregative Escherichia coli and its virulence-related genes in a case-control study among children from north-eastern Brazil. J. Med. Microbiol. 62, 683–693. doi: 10.1099/jmm.0.054262-0
Liu, B., Zheng, D., Jin, Q., Chen, L., and Yang, J. (2019). VFDB 2019: a comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 47, D687–D692. doi: 10.1093/nar/gky1080
Liu, M., Li, X., Xie, Y., Bi, D., Sun, J., Li, J., et al. (2019). ICEberg 2.0: an updated database of bacterial integrative and conjugative elements. Nucleic Acids Res. 47, D660–D665. doi: 10.1093/nar/gky1123
Maddison, W. P., and Maddison, D. R. (2019). Mesquite: a Modular System for Evolutionary Analysis. Version 3.61. Available online at: http://www.mesquiteproject.org
Maldonado-Contreras, A., Birtley, J. R., Boll, E., Zhao, Y., Mumy, K. L., Toscano, J., et al. (2017). Shigella depends on SepA to destabilize the intestinal epithelial integrity via cofilin activation. Gut Microbes 8, 544–560. doi: 10.1080/19490976.2017.1339006
Manning, S. D., Motiwala, A. S., Springman, A. C., Qi, W., Lacher, D. W., Ouellette, L. M., et al. (2008). Variation in virulence among clades of Escherichia coli O157:H7 associated with disease outbreaks. Proc. Natl. Acad. Sci. U.S.A. 105, 4868–4873. doi: 10.1073/pnas.0710834105
Mohawk, K. L., Melton-Celsa, A. R., Zangari, T., Carroll, E. E., and O'brien, A. D. (2010). Pathogenesis of Escherichia coli O157:H7 strain 86-24 following oral infection of BALB/c mice with an intact commensal flora. Microb. Pathog. 48, 131–142. doi: 10.1016/j.micpath.2010.01.003
Morelli, G., Song, Y., Mazzoni, C. J., Eppinger, M., Roumagnac, P., Wagner, D. M., et al. (2010). Yersinia pestis genome sequencing identifies patterns of global phylogenetic diversity. Nat. Genet. 42, 1140–1143. doi: 10.1038/ng.705
Morin, N., Santiago, A. E., Ernst, R. K., Guillot, S. J., and Nataro, J. P. (2013). Characterization of the AggR regulon in enteroaggregative Escherichia coli. Infect. Immun. 81, 122–132. doi: 10.1128/IAI.00676-12
Myers, G. S., Mathews, S. A., Eppinger, M., Mitchell, C., O'brien, K. K., White, O. R., et al. (2009). Evidence that human Chlamydia pneumoniae was zoonotically acquired. J. Bacteriol. 191, 7225–7233. doi: 10.1128/JB.00746-09
Nataro, J. P., Kaper, J. B., Robins-Browne, R., Prado, V., Vial, P., and Levine, M. M. (1987). Patterns of adherence of diarrheagenic Escherichia coli to HEp-2 cells. Pediatr. Infect. Dis. J. 6, 829–831. doi: 10.1097/00006454-198709000-00008
Okeke, I. N., and Nataro, J. P. (2001). Enteroaggregative Escherichia coli. Lancet Infect. Dis. 1, 304–313. doi: 10.1016/S1473-3099(01)00144-X
Olson, S., Hall, A., Riddle, M. S., and Porter, C. K. (2019). Travelers' diarrhea: update on the incidence, etiology and risk in military and similar populations - 1990-2005 versus 2005-2015, does a decade make a difference? Trop. Dis. Travel Med. Vaccines 5:1. doi: 10.1186/s40794-018-0077-1
Pichel, M., Binsztein, N., and Viboud, G. (2000). CS22, a novel human enterotoxigenic Escherichia coli adhesin, is related to CS15. Infect. Immun. 68, 3280–3285. doi: 10.1128/IAI.68.6.3280-3285.2000
Porter, C. K., Olson, S., Hall, A., and Riddle, M. S. (2017). Travelers' diarrhea: an update on the incidence, etiology, and risk in military deployments and similar travel populations. Mil. Med. 182, 4–10. doi: 10.7205/MILMED-D-17-00064
Riddle, M. S., Connor, P., Fraser, J., Porter, C. K., Swierczewski, B., Hutley, E. J., et al. (2017). Trial evaluating ambulatory therapy of travelers' diarrhea (TrEAT TD) study: a randomized controlled trial comparing 3 single-dose antibiotic regimens with loperamide. Clin. Infect. Dis. 65, 2008–2017. doi: 10.1093/cid/cix693
Ross, B. N., Rojas-Lopez, M., Cieza, R. J., Mcwilliams, B. D., and Torres, A. G. (2015). The role of long polar fimbriae in Escherichia coli O104:H4 adhesion and colonization. PLoS ONE 10:e0141845. doi: 10.1371/journal.pone.0141845
Roux, A., Beloin, C., and Ghigo, J. M. (2005). Combined inactivation and expression strategy to study gene function under physiological conditions: application to identification of new Escherichia coli adhesins. J. Bacteriol. 187, 1001–1013. doi: 10.1128/JB.187.3.1001-1013.2005
Ruiz-Perez, F., and Nataro, J. P. (2014). Bacterial serine proteases secreted by the autotransporter pathway: classification, specificity, and role in virulence. Cell. Mol. Life Sci. 71, 745–770. doi: 10.1007/s00018-013-1355-8
Rusconi, B., Sanjar, F., Koenig, S. S., Mammel, M. K., Tarr, P. I., and Eppinger, M. (2016). Whole genome sequencing for genomics-guided investigations of Escherichia coli O157:H7 outbreaks. Front. Microbiol. 7:985. doi: 10.3389/fmicb.2016.00985
Santiago, A. E., Ruiz-Perez, F., Jo, N. Y., Vijayakumar, V., Gong, M. Q., and Nataro, J. P. (2014). A large family of antivirulence regulators modulates the effects of transcriptional activators in Gram-negative pathogenic bacteria. PLoS Pathog. 10:e1004153. doi: 10.1371/journal.ppat.1004153
Sheikh, J., Czeczulin, J. R., Harrington, S., Hicks, S., Henderson, I. R., Le Bouguenec, C., et al. (2002). A novel dispersin protein in enteroaggregative Escherichia coli. J. Clin. Invest. 110, 1329–1337. doi: 10.1172/JCI16172
Sheikh, J., Dudley, E. G., Sui, B., Tamboura, B., Suleman, A., and Nataro, J. P. (2006). EilA, a HilA-like regulator in enteroaggregative Escherichia coli. Mol. Microbiol. 61, 338–350. doi: 10.1111/j.1365-2958.2006.05234.x
Sheikh, J., Hicks, S., Dall'agnol, M., Phillips, A. D., and Nataro, J. P. (2001). Roles for fis and yafk in biofilm formation by enteroaggregative Escherichia coli. Mol. Microbiol. 41, 983–997. doi: 10.1046/j.1365-2958.2001.02512.x
Siguier, P., Perochon, J., Lestrade, L., Mahillon, J., and Chandler, M. (2006). ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 34, D32–D36. doi: 10.1093/nar/gkj014
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Subramanian, B., Gao, S., Lercher, M. J., Hu, S., and Chen, W. H. (2019). Evolview v3: a webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 47, W270–W275. doi: 10.1093/nar/gkz357
Wilgenbusch, J. C., and Swofford, D. (2003). Inferring evolutionary trees with PAUP*. Curr. Protoc. Bioinformatics Chapter 6: Unit 6.4. doi: 10.1002/0471250953.bi0604s00
Xie, Z., and Tang, H. (2017). ISEScan: automated identification of insertion sequence elements in prokaryotic genomes. Bioinformatics 33, 3340–3347. doi: 10.1093/bioinformatics/btx433
Zaidi, D., and Wine, E. (2015). An update on travelers' diarrhea. Curr. Opin. Gastroenterol. 31, 7–13. doi: 10.1097/MOG.0000000000000133
Zangari, T., Melton-Celsa, A. R., Panda, A., Boisen, N., Smith, M. A., Tatarov, I., et al. (2013). Virulence of the Shiga toxin type 2-expressing Escherichia coli O104:H4 German outbreak isolate in two animal models. Infect. Immun. 81, 1562–1574. doi: 10.1128/IAI.01310-12
Zankari, E., Hasman, H., Cosentino, S., Vestergaard, M., Rasmussen, S., Lund, O., et al. (2012). Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 67, 2640–2644. doi: 10.1093/jac/dks261
Zboromyrska, Y., Hurtado, J. C., Salvador, P., Alvarez-Martinez, M. J., Valls, M. E., Mas, J., et al. (2014). Aetiology of traveller's diarrhoea: evaluation of a multiplex PCR tool to detect different enteropathogens. Clin. Microbiol. Infect. 20, O753–O759. doi: 10.1111/1469-0691.12621
Keywords: enteroaggregative, Escherichia coli, travelers' diarrhea, EAEC, mouse model, biofilm, aap, deployed military personnel
Citation: Petro CD, Duncan JK, Seldina YI, Allué-Guardia A, Eppinger M, Riddle MS, Tribble DR, Johnson RC, Dalgard CL, Sukumar G, Connor P, Boisen N and Melton-Celsa AR (2020) Genetic and Virulence Profiles of Enteroaggregative Escherichia coli (EAEC) Isolated From Deployed Military Personnel (DMP) With Travelers' Diarrhea. Front. Cell. Infect. Microbiol. 10:200. doi: 10.3389/fcimb.2020.00200
Received: 02 December 2019; Accepted: 16 April 2020;
Published: 20 May 2020.
Edited by:
Alexandra Aubry, Sorbonne Universités, FranceReviewed by:
Waldir P. Elias, Butantan Institute, BrazilCopyright © 2020 Petro, Duncan, Seldina, Allué-Guardia, Eppinger, Riddle, Tribble, Johnson, Dalgard, Sukumar, Connor, Boisen and Melton-Celsa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Angela R. Melton-Celsa, YW5nZWxhLm1lbHRvbi1jZWxzYUB1c3Vocy5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.