 Regiane C. B. Dias1
Regiane C. B. Dias1 Rodrigo H. S. Tanabe1
Rodrigo H. S. Tanabe1 Melissa A. Vieira1
Melissa A. Vieira1 Maria C. Cergole-Novella2
Maria C. Cergole-Novella2 Luís F. dos Santos3
Luís F. dos Santos3 Tânia A. T. Gomes4
Tânia A. T. Gomes4 Waldir P. Elias5
Waldir P. Elias5 Rodrigo T. Hernandes1*
Rodrigo T. Hernandes1*- 1Universidade Estadual Paulista (UNESP), Instituto de Biociências, São Paulo, Brazil
- 2Laboratório Regional de Santo André, Instituto Adolfo Lutz, São Paulo, Brazil
- 3Centro de Bacteriologia, Instituto Adolfo Lutz, São Paulo, Brazil
- 4Departamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal de São Paulo- Escola Paulista de Medicina (UNIFESP-EPM), São Paulo, Brazil
- 5Laboratório de Bacteriologia, Instituto Butantan, São Paulo, Brazil
Enteroaggregative Escherichia coli (EAEC) is an important agent of acute and persistent diarrhea in children and adults worldwide. Here we report a characterization of 220 EAEC isolates, 88.2% (194/220) of which were typical and 11.8% (26/220) were atypical, obtained from diarrheal patients during seven years (2010-2016) of epidemiological surveillance in Brazil. The majority of the isolates were assigned to phylogroups A (44.1%, 97/220) or B1 (21.4%, 47/220). The aggregative adherence (AA) pattern was detected in 92.7% (204/220) of the isolates, with six of them exhibiting AA concomitantly with a chain-like adherence pattern; and agg5A and agg4A were the most common adhesin-encoding genes, which were equally detected in 14.5% (32/220) of the isolates. Each of 12 virulence factor-encoding genes (agg4A, agg5A, pic, aap, aaiA, aaiC, aaiG, orf3, aar, air, capU, and shf) were statistically associated with typical EAEC (P < 0.05). The genes encoding the newly described aggregate-forming pili (AFP) searched (afpB, afpD, afpP, and afpA2), and/or its regulator (afpR), were exclusively detected in atypical EAEC (57.7%, 15/26), and showed a significant association with this subgroup of EAEC (P < 0.001). In conclusion, we presented an extensive characterization of the EAEC circulating in the Brazilian settings and identified the afp genes as putative markers for increasing the efficiency of atypical EAEC diagnosis.
Introduction
Enteroaggregative Escherichia coli (EAEC) is one of the six diarrheagenic E. coli (DEC) pathotypes, which is defined as E. coli isolates that produce the aggregative adherence pattern (AA) on cultured epithelial cells (HeLa and/or HEp-2) in vitro (Nataro and Kaper, 1998; Kaper et al., 2004; Croxen et al., 2013). The AA pattern is characterized by a bacterial arrangement that resembles stacked bricks on both epithelial cells and the coverslip (Nataro et al., 1987). EAEC is a cause of diarrheal disease in children and adults, living in both developing and developed countries, and are associated with cases of persistent diarrhea and diarrheal outbreaks worldwide (Huang et al., 2006; Estrada-Garcia and Navarro-Garcia, 2012; Hebbelstrup Jensen et al., 2014), besides the fact that EAEC can also be isolated from asymptomatic subjects (Gioppo et al., 2000; Araujo et al., 2007; Lima et al., 2013, 2019a; Dias et al., 2016; Durand et al., 2016).
Previous studies have shown that EAEC isolates can be identified in several serotypes (Zamboni et al., 2004; Boisen et al., 2012; Lima et al., 2019b), and are distributed across the four main E. coli phylogroups (A, B1, B2, and D) (França et al., 2013; Imuta et al., 2016). Moreover, high rates of resistance to ampicillin and trimethoprim-sulphamethoxazole, but not to quinolones, have been observed (Aslani et al., 2011; Ikumapayi et al., 2017).
The pathogenicity of EAEC is characterized by the colonization of the intestinal epithelium, followed by the production and secretion of enterotoxins and cytotoxins, which result in secretory diarrhea (Nataro and Kaper, 1998). The establishment of the AA pattern has been associated with the presence of five distinct aggregative adherence fimbriae (AAF/I-AAF/V) (Nataro et al., 1992; Czeczulin et al., 1997; Bernier et al., 2002; Boisen et al., 2008; Jønsson et al., 2015), which belong to the chaperone-usher subclass, as well as afimbrial adhesins (Monteiro-Neto et al., 2003; Bhargava et al., 2009). Very recently, a type IV pilus, termed aggregate-forming pili (AFP), encoded by genes located in the afp operon, was identified to be responsible for the establishment of the AA pattern by a hybrid EAEC/STEC (Shiga-toxin producing E. coli) isolate of serotype O23:H8, that lacks the AAFs described so far (Lang et al., 2018).
Once adhered to the intestinal epithelium, EAEC produces several enterotoxins and cytotoxins, which include: the enteroaggregative E. coli heat-stable enterotoxin (EAST-1), encoded by the astA gene, the plasmid-encoded toxin (Pet), the protein involved in colonization (Pic), and autotransporter proteases such as SigA and SepA, which were initially described in Shigella flexneri. Another important virulence factor associated with the pathogenicity of EAEC is the protein dispersin (Aap), which facilitates bacterial dispersion in the intestinal epithelium (Sheikh et al., 2002). Dispersin secretion occurs via an ATP-binding cassette (ABC) transporter system, encoded by a cluster of five genes (aatPABCD) located in the aggregative adherence plasmid (pAA). The aatA gene, which corresponds to the former EAEC probe CVD432, is one the most used target for EAEC identification nowadays (Baudry et al., 1990; Nishi et al., 2003; Lozer et al., 2013).
The AggR protein, encoded by the aggR gene located in the pAA, is a global virulence regulator, which activates the expression of several plasmidial and chromosomal virulence factor-encoding genes in the EAEC 042 prototype strain, such as the AAFs, as well as a chromosomal pathogenicity island (PAI), termed aaiA-aaiP, which encodes a Type VI Secretion System (T6SS) (Dudley et al., 2006; Morin et al., 2013). Some studies employed genes of the aaiA-aaiP operon, such as aaiA, aaiC, and aaiG, as an additional chromosomal marker to enhance the efficiency of EAEC diagnosis (Lima et al., 2013; Andrade et al., 2014; Havt et al., 2017; Hebbelstrup Jensen et al., 2018). Based on the presence of the aggR gene, EAEC isolates can be subgrouped into typical (aggR+) and atypical (aggR−) (Kaper et al., 2004).
Moreover, other potential virulence factor-encoding genes that may contribute to the pathogenicity of EAEC isolates, such as aar (AggR-activated regulator), shf (which encodes a protein involved in biofilm formation initially identified in Shigella flexneri), air (Enteroaggregative immunoglobulin repeat protein), and capU (which encodes a homologous hexosyltransferase) have been described (Czeczulin et al., 1999; Sheikh et al., 2006; Santiago et al., 2017).
The goal of this study was to better understand the diversity of typical and atypical EAEC isolates, obtained from diarrheal patients in Brazil, regarding the somatic (O) and flagellar (H) antigens determination (serotypes), phylogenetic classification, presence of 25 virulence factors-encoding genes (e.g., adhesins, toxins, secreted proteases, type VI secretion system), adherence pattern produced on HeLa cells, and the susceptibility to 11 distinct antimicrobial drugs.
Materials and Methods
Enteroaggregative Escherichia coli (EAEC) Isolates Used in This Study
A total of 220 EAEC isolates, obtained from stool samples of sporadic cases of diarrhea occurred in Brazil, were included in this study. Of note, 203 EAEC isolates were obtained over 6-years of active epidemiological surveillance (2011–2016), performed at the Instituto Adolfo Lutz (IAL), a public health laboratory and the Brazilian reference center for diarrheagenic E. coli identification, as previously published (Ori et al., 2019), as well as 17 EAEC isolates, obtained during the year of 2010, at the same Institute.
In this study, EAEC was defined as E. coli isolates that possess the aatA gene (former CVD432 probe), and typical and atypical EAEC were differentiated based on the presence of the aggR gene in the former group (Kaper et al., 2004).
Serotyping
The somatic (O) and flagellar (H) antigens from 203 EAEC isolates, obtained from 2011 to 2016, were determined in our previous epidemiological surveillance study (Ori et al., 2019). In the 17 remaining EAEC isolates, obtained during the year of 2010, the O and H antigens were determined by standard agglutination tests, with O (O1–O181) and H (H1–H56) antisera produced at Instituto Adolfo Lutz, São Paulo, Brazil (Ewing, 1986).
Phylogroup classification
The classification of the 220 EAEC isolates studied into distinct phylogroups (A, B1, B2, C, D, E, and F) and Escherichia clades were performed using the quadruplex Polymerase Chain Reaction (PCR) method as previously described by Clermont et al. (2013). Subsequently, EAEC isolates classified in the phylogroup B2, with the following genotype: arpA−, chuA+, yjaA−, and TspE4+, and all isolates assigned in the phylogroup F were tested in an additional Triplex PCR, using primers and PCR conditions as recently described (Clermont et al., 2019), in order to confirm these isolates as B2 and F or to reclassify them in the phylogroup G.
Detection of Virulence Factor-Encoding Genes
The presence or absence of the EAEC diagnosis markers aatA and aggR was confirmed in all 220 isolates, and subsequently, these isolates were tested for the presence of genes encoding for: adhesins (aggA, aafA, agg3A, agg4A, agg5A, afpB, afpD, afpP, afpA2, afpR, and eibG), toxins (pic, pet, astA, sigA, and sepA) and other virulence factors (aap, aaiA, aaiC, aaiG, orf3, aar, air, capU, and shf), by PCR DNA amplification. PCR was performed using GoTaq Green Master Mix (Promega, Madison, WI, USA) with 0.34 μM of each of the primers. All primer sequences and PCR assay conditions used for the detection of virulence genes are described in the references cited in Table S1. PCR products were subjected to agarose gel electrophoresis, prepared with Tris-borate-EDTA (TBE) buffer, and visualized with SYBR Safe DNA Gel Stain (Invitrogen, CA, USA).
Antimicrobial Susceptibility Test
Antimicrobial susceptibility assays were performed following the recommendations of the Clinical and Laboratory Standards Institute (CLSI, 2018), using the disk diffusion method on Mueller-Hinton agar (OXOID, UK). The antimicrobial drugs tested were: ampicillin (AMP; 10 μg), amoxicillin-clavulanic acid (AMC; 20/10 μg), cefazolin (CFZ; 30 μg), cefuroxime (CFX; 30 μg), cefotaxime (CTX; 30 μg), cefepime (CPM; 30 μg), imipenem (IPM; 10 μg), gentamicin (GEN; 10 μg), tobramycin (TOB; 10 μg), ciprofloxacin (CIP; 5 μg), and trimetoprim-sulfamethoxazole (SUT, 1.25/23.75 μg). The E. coli ATCC 25922 was used as quality control, and the commercial disks were obtained from Cefar Diagnóstica Ltda (SP, Brazil). EAEC multidrug-resistant was defined by the detection of isolates resistant to three or more classes of antibiotics tested (Magiorakos et al., 2012; Chattaway et al., 2017).
Adherence Assays
The adherence patterns of the EAEC isolates was evaluated in HeLa cells as previously described (Cravioto et al., 1979), with some modifications. Cells were cultured in DMEM (Dulbecco's Modified Eagle Medium—high glucose; Sigma-Aldrich; St. Louis, MO, USA), supplemented with 10% fetal bovine serum (Gibco; Grand Island, New York, USA), with 1% antibiotic (Penicillin-Streptomycin, Sigma; St. Louis, MO, USA) in 5% CO2 at 37°C. For the adherence assay, ~1 × 105 cells were cultivated in a 24-well microplate, using the same medium described above and containing glass coverslips in each well of the microplate, for 48 h until reaching a confluence of ~80%. Then, the cells were washed three times with phosphate-buffered saline (PBS, pH 7.4), 1.0 mL of DMEM, supplemented with 2% fetal bovine serum and 1% D-mannose (Sigma, Saint Louis, MO, USA) was added, the cells were infected with 20 μL of an overnight bacterial suspension in LB broth (containing ~108 CFU/mL), and the adherence assay was performed in 5% CO2 at 37°C. After 3 h of incubation, the infected cells were washed six times with PBS and then fixed with methanol (Dinâmica, SP, Brazil) and stained with May-Grünwald (Sigma; St. Louis, MO, USA) and Giemsa (Dinâmica, SP, Brazil) for identification of the adherence pattern by light microscopy analyzes. When the EAEC isolates did not adhere to the epithelial cells or the adherence pattern could not be defined, the adherence assay was repeated in a 6 h bacteria-cell interaction assay, with a washing step after 3 h of incubation, as previously described (Rodrigues et al., 1996), stained with May-Grünwald/Giemsa and then observed by light microscopy. To eliminate potential bias in the adherence pattern classification, each slide was analyzed by three distinct blinded examiners, both to define the adherence pattern exhibited by each EAEC isolate, as well as to select the representative images to be included in Figures 1, 2. In cases of divergence among the three examiners, the EAEC isolate was submitted to a new adherence assay and reevaluated in a blinded-manner until a consensus was obtained.
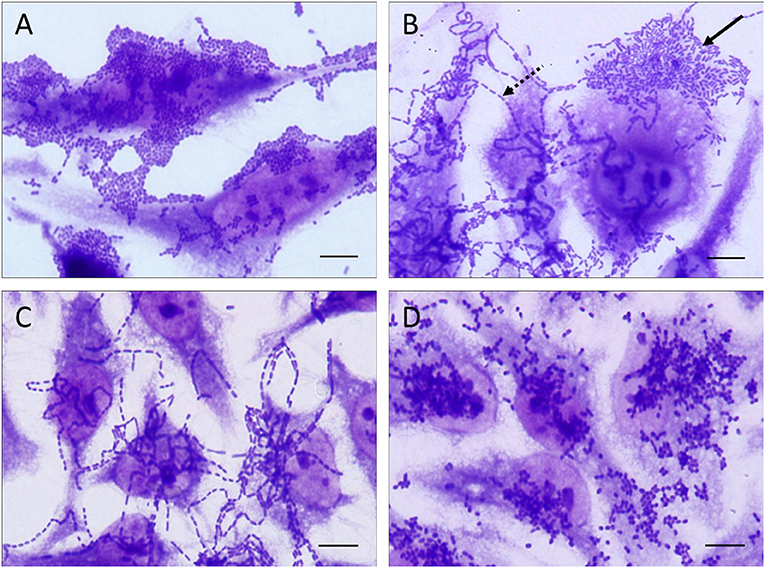
Figure 1. Representative images of the distinct patterns of adherence exhibited by the EAEC isolates studied in adherence assays performed with HeLa cells. The adherence patterns are illustrated as follow: (A) aggregative adherence pattern, (B) aggregative adherence pattern (solid arrow) occurring concomitantly with the chain-like adherence (dashed arrow), (C) chain-like adherence pattern, and (D) diffuse adherence pattern. All the adherence patterns illustrated in this picture were observed in adherence assays performed with 3 h of infection. Scale bar = 10 μm.
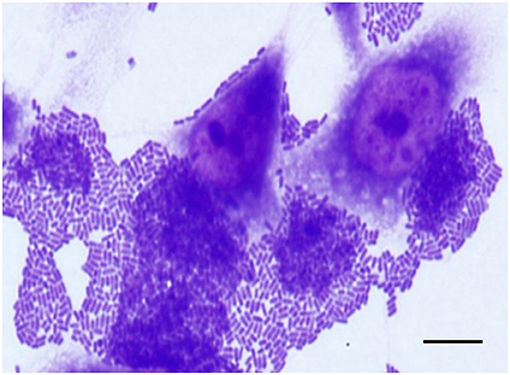
Figure 2. Aggregative adherence (AA) pattern exhibited by an atypical EAEC (aatA+/aggR−) isolate (IAL 6006) that harbored the afp operon. Note the bacterial stacked brick-like arrangement, characteristic of the AA pattern, on both epithelial cells and coverslip surfaces detected in adherence assay performed with 3 h of infection. Scale bar = 10 μm.
Statistical Analyses
Differences observed between typical and atypical EAEC were investigated using a Chi-square with Yates' correction or two-tailed Fisher's exact test, as appropriate, and a significance level of 5% (P < 0.05) was adopted to consider the differences statistically significant.
Results
The majority of the diarrheal patients, from which the EAEC isolates were obtained, were children up to five-years-old (62.7%, 138/220) and from the São Paulo State (66.8%, 147/220), besides EAEC isolates that were obtained from other Brazilian States, such as Minas Gerais (18.6%, 41/220) and Santa Catarina (14.5%, 32/220) (Table S2). We detected 194 typical (88.2%, 194/220) and 26 (11.8%, 26/220) atypical EAEC (Table 1), and observed that 56.8% (125/220) were classified in 39 distinct serogroups, besides the occurrence of O non-typeable (19.5%, 43/220) and rough (23.6%, 52/220) isolates (Table S3). Regarding the serotypes, O15:H2 (4.5%; 10/220), O175:H28 (4.5%; 10/220), O73:H1 (4.1%; 9/220), O153:H2 (3.6%; 8/220), and O21:H2 (3.2%; 7/220) were the most frequent ones, and exclusively detected in the typical EAEC subgroup (Table S3). The classification of the EAEC isolates in the distinct E. coli phylogroups, demonstrated that EAEC were assigned mainly in the phylogroups A (44.1%, 97/220), B1 (21.4%, 47/220), D (13.2%, 29/220), and E (10.0%, 22/220) (Table 2), and the number of atypical EAEC identified in the phylogroup A was significantly higher than typical (65.4 vs. 41.2%), with this difference being statistically significant (P < 0.05). Moreover, 2.3% (5/220) of the EAEC isolates studied, serotypes O73:H1 (3 isolates) and O15:H18 (2 isolates), were identified in the newly described phylogroup G (Table 2).
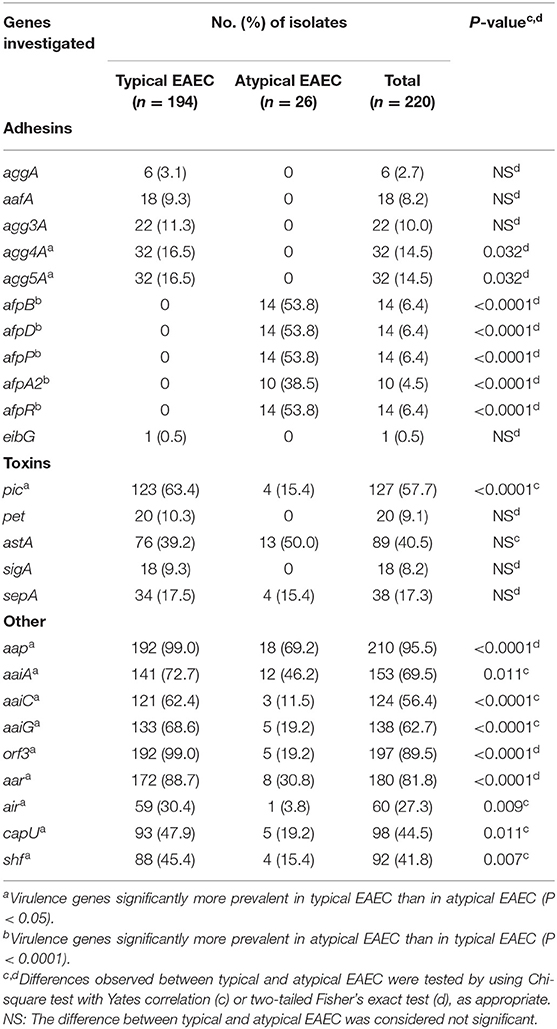
Table 1. Occurrence of virulence factor-encoding genes in typical and atypical EAEC isolates.
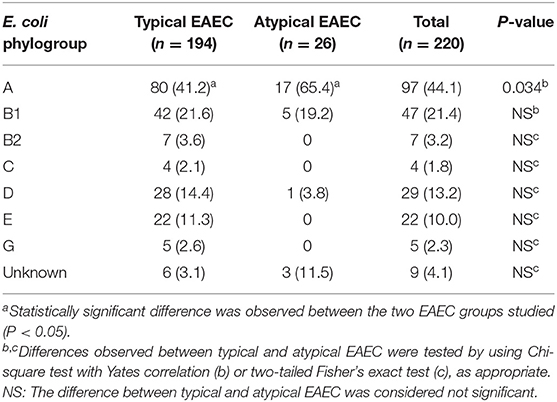
Table 2. Classification of the typical and atypical EAEC isolates in the distinct E. coli phylogroups.
We also investigated the occurrence of 25 virulence genes encoding adhesins, toxins, secreted proteases, and T6SS (Table 1). Genes encoding the major subunit of the five distinct AAFs were exclusively detected in typical EAEC, with agg4A (16.5%, 32/194) and agg5A (16.5%, 32/194) being the most frequent ones (Table 1). Interestingly, genes encoding proteins associated with the biogenesis of the newly described AFP adhesin (afpB, afpD, afpP, and afpA2) and/or its regulator (afpR) were exclusively detected in atypical EAEC (57.7%, 15/26), showing a statistical association with this subgroup (P < 0.0001). Typical EAEC harbored all five genes encoding for toxins investigated in this study, such as pic, pet, astA, sigA, and sepA, while atypical EAEC isolates lacked the pet and sigA genes. Furthermore, pic was significantly more frequent in typical than atypical EAEC (63.4 vs. 15.4%, P < 0.0001). Additionally, the genes encoding for dispersin (aap), T6SS (aaiA, aaiC, and aaiG) and five other putative virulence factors (orf3, aar, air, capU, and shf) were detected in typical and atypical, despite their statistical association with the former group (Table 1). The genes encoding the T6SS investigated here (aaiA, aaiC, and aaiG) were detected in 69.5% (153/220) of the EAEC isolates studied, with the concomitant occurrence of the three genes being the most common genotype (55.5%, 122/220) detected (Table 3). Besides the aforementioned combination, the aaiA gene was observed in association with aaiG (7.3%, 16/220), aaiC (0.9%, 2/220), or not associated with these two genes (5.9%, 13/220) (Table 3).
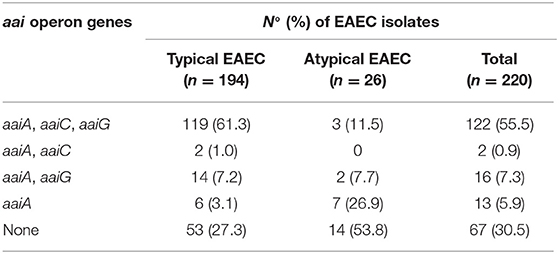
Table 3. Occurrence of the aai operon genes in typical and atypical EAEC isolates.
The AA pattern was detected in 92.7% (204/220) of the isolates (Table 4). In 75.9% (167/220) of them, the AA pattern was detected after 3 h of bacteria-cell interaction (Figure 1A), while in 14.1% (31/220) this phenotype was detected using more prolonged periods of incubation (6 h) (Table 4). Curiously, six EAEC isolates (2.7%, 6/220) produced AA concomitantly with long-chain aggregates of bacteria adhered to both HeLa cell surfaces and coverslip (Figure 1B), which is characteristic of an adherence pattern termed chain-like adhesion (CLA). We also observed that 1.4% (3/220) and 0.9% (2/220) of the EAEC isolates produced CLA (Figure 1C) or the diffuse adherence (DA) patterns (Figure 1D), respectively. EAEC isolates exhibiting an undefined (UND) adherence (1.4%, 3/220) or unable to adhere to HeLa cells (3.6%, 8/220), even using more prolonged periods of bacteria-cell interaction, were observed (Table 4). The majority of the typical EAEC isolates (82.4%, 89/108) that harbored one or more of the adhesin-encoding genes investigated were able to produce the AA pattern within 3 h of assay, and the only EAEC that harbored the eibG gene produced CLA (Table 4). Interestingly, two typical EAEC isolates harbored the agg3A concomitantly with the agg5A, and both the isolates were able to produce the AA pattern (Table 4). Among the 26 atypical EAEC, 76.9% (20/26) produced AA, besides isolates exhibiting UND adherence (7.7%, 2/26), or non-adherent (NA) (15.4%, 4/26) (Table 4). Taken into consideration the 14 afp+ EAEC isolates, 85.7% (12/14) produced AA, while the other two isolates (14.3%, 2/14) were NA (Table 4). The isolate IAL 5749 that harbored only the regulator-encoding gene (afpR), but lacked the genes from the afp operon investigated, produced a UND adherence pattern to HeLa cells (Table 4).
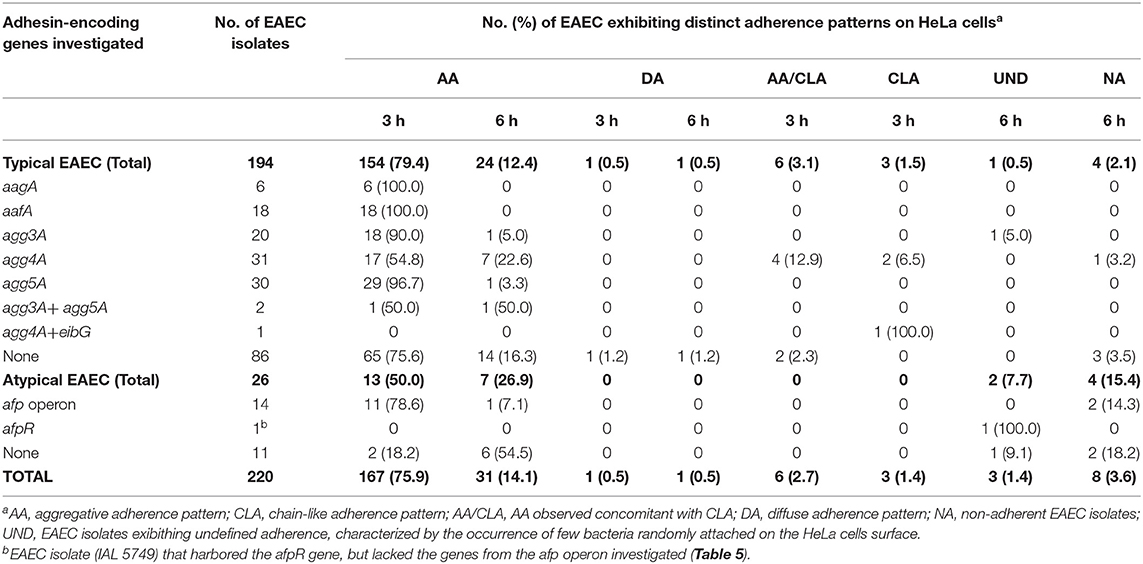
Table 4. Correlation of adhesin-encoding genes and adherence patterns detected in the typical and atypical EAEC isolates studied.
Considering the novelty of the association of the genes of the afp operon and/or its regulator (afpR) with atypical EAEC, we compiled the main features of these 15 EAEC isolates in Table 5. Most of them were identified in the phylogroup A (80.0%, 12/15) and produced AA in 3 h assay (73.3%, 11/15) (Figure 2), even though they were heterogeneous regarding the serotype, and virulence profile exhibited (Table 5).
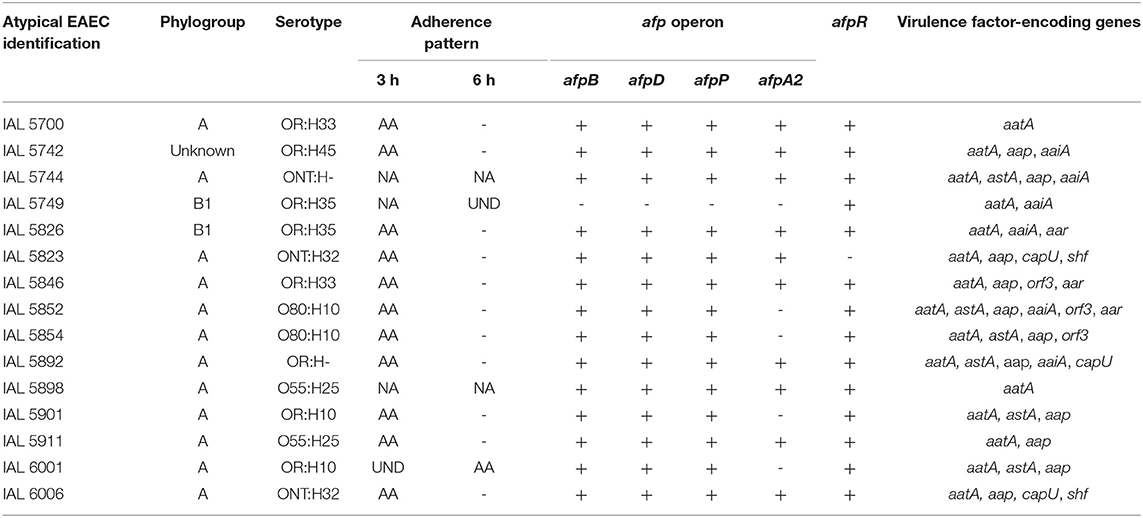
Table 5. Phenotypic and molecular features of atypical EAEC harboring the afp operon gene.
Among the 11 antimicrobial drugs tested, non-susceptible typical EAEC isolates were observed for ampicillin, amoxicillin/clavulanic acid, cefazolin, gentamicin, tobramycin, ciprofloxacin, and trimethoprim/sulfamethoxazole, with the highest resistance rates observed for ampicillin (52.6%, 102/194), and trimethoprim/sulfamethoxazole (33.5%, 65/194) (Table 6). Non-susceptible atypical EAEC were observed only for ampicillin (19.2%, 5/26) and trimethoprim/sulfamethoxazole (19.2%, 5/26) (Table 6). Multidrug resistance phenotype was observed only in two (1.0%, 2/194) of the typical and in none of the atypical EAEC isolates studied (Table S4).
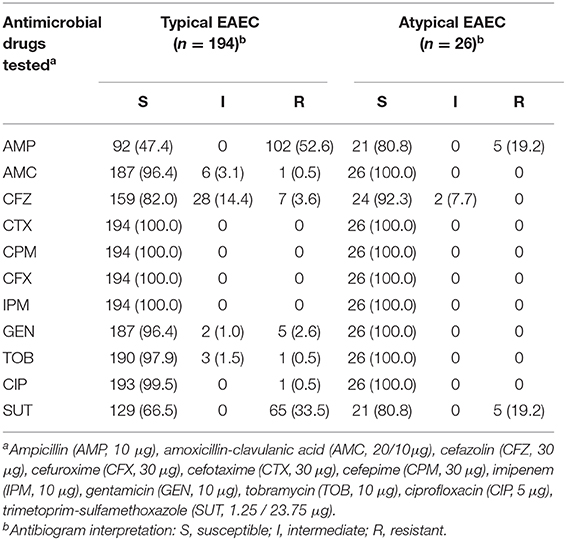
Table 6. Antimicrobial susceptibility of the EAEC isolates studied.
Discussion
The importance of EAEC as an etiological agent of diarrheal diseases has been reinforced in the last decade (Boisen et al., 2012; Lima et al., 2013; Durand et al., 2016; Chattaway et al., 2017; Ori et al., 2019). However, some questions remain to be addressed, such as the use of the adherence pattern as the gold standard for diagnosis, how to identify truly pathogenic isolates in this heterogeneous group of isolates, and which would be the best molecular markers to improve the diagnosis of EAEC, mainly atypical EAEC.
Several of the virulence factor-encoding genes investigated here were statistically more frequent among typical EAEC, which is not surprising since the majority of these genes were identified, and functionally characterized, in the typical EAEC prototype strain 042 (Harrington et al., 2006; Chaudhuri et al., 2010; Hebbelstrup Jensen et al., 2014). Similar findings were also observed by other Brazilian authors that compared typical and atypical EAEC isolates, especially with the pic, aap, shf, and agg4A genes being more frequent among the typical EAEC isolates (Andrade et al., 2017; Taborda et al., 2018; Guerrieri et al., 2019).
Case-control studies, performed in distinct geographic regions, have been able to demonstrate the association of some genes encoding virulence factors with diarrheal disease. pic, which encodes a protein with hemagglutinin and mucinolytic activity in vitro (Henderson et al., 1999), was statistically more frequent among diarrheal Peruvian children than in healthy subjects (Durand et al., 2016), as well as sepA, encoding an autotransporter protease originally described in Shigella spp., was associated with diarrhea in children from Bamako, Mali (Boisen et al., 2012). A case-control study performed in Northeastern Brazil demonstrated the association of pet (Plasmid-encoded toxin) and aafA (AAF/II fimbrial subunit) with diarrhea (Lima et al., 2013). Moreover, the occurrence of typical EAEC, harboring concomitantly aatA and aaiC, was more frequent among malnourished than in nourished children (Havt et al., 2017), thus reinforcing the need for greater attention to asymptomatic infections by this pathogen.
In typical EAEC the establishment of the AA pattern is mediated by five distinct AAFs (Jønsson et al., 2017a), and the occurrence of genes associated with the biogenesis of AAF/I-AAF/IV has already been reported among Brazilian EAEC isolates, with their frequency varying according to the geographic regions studied (Elias et al., 1999; Zamboni et al., 2004; Lima et al., 2013; Guerrieri et al., 2019). However, the occurrence of Brazilian EAEC isolates harboring the agg5A, alone or concomitantly with agg3A, is described for the first time in this study. A study performed with Danish EAEC isolates identified six harboring the major pilin subunits from both AAF/III (agg3A) and AAF/V (agg5A), with these genes located in a novel pAA variant (Jønsson et al., 2017b).
Recently, Lang and coworkers described a new fimbria, termed aggregate-forming pili (AFP), in a hybrid EAEC/STEC isolate of serotype O23:H8 (Lang et al., 2018). The genes responsible for the biogenesis of AFP have ~52% of nucleotide identity with the genes responsible for encoding the bundle-forming pili (BFP), a type IV pilus that mediates the localized adherence (LA) phenotype of typical enteropathogenic E. coli on epithelial cells surface (Scaletsky et al., 1984; Girón et al., 1991; Donnenberg et al., 1992). The afp and bfp operons have a similar gene organization (Tobe et al., 1999; Lang et al., 2018), despite the presence of an additional afpA2 gene, that is not required for AFP biogenesis (Lang et al., 2018). The deletion of the complete afp operon, afpA and afpR genes (encoding an AraC-like regulator), but not afpA2, led to the loss of fimbriae production and drastically reduced the ability of the EAEC/STEC isolate in adhering to epithelial cells in vitro (Lang et al., 2018).
Using an NCBI nucleotide BLAST analysis with the afp operon genes as a query sequence, Lang et al. (2018) identified 17 afp-positive E. coli isolates, which were also positive for EAEC markers, such as aatA and aap, but lacking aggR, which is a genotype of atypical EAEC. This observation motived us to deeply investigate the occurrence of the afp genes in other EAEC isolates, including both typical and atypical. To the best of our knowledge, this is the first study to demonstrate the association of genes from the afp operon, as well as, a gene encoding its regulator (afpR) with the atypical EAEC subgroup, thus providing a new putative molecular marker to increase the diagnostic efficiency of this pathogen.
Moreover, some of the isolates classified as typical EAEC (aatA+/aggR+) produced the CLA or DA patterns on HeLa cells (3 and 2 isolates, respectively), similar to the observed in previous studies that compared adherence pattern and molecular markers of the distinct DEC pathotypes (Gomes et al., 1998; Gioppo et al., 2000; Scaletsky et al., 2002; Spano et al., 2017). A Brazilian study demonstrated that E. coli isolates exhibiting CLA on epithelial cells harbored genes commonly used as a marker of the EAEC pathotype, i.e., aatA and aggR, as well as virulence factor-encoding genes, such as pet, astA, and aggC (Gioppo et al., 2000). These data demonstrate that the EAEC definition is still ambiguous since E. coli isolates producing the AA pattern on HeLa cells could be devoid of EAEC molecular markers of this pathotype, as well as E. coli positive for such genes could produce adherence patterns distinct from AA. We believe that further studies using whole-genome sequencing analysis, in combination with phenotypic features of the EAEC isolates and clinical information of the patients could provide greater accuracy to the molecular markers necessary for the EAEC diagnosis, as well as, in the identification of truly pathogenic isolates.
In conclusion, although we have characterized the EAEC isolates studied in terms of phylogroups, serotypes, adherence patterns, and genes that encode virulence factors, the main message of this study is the association of the aggregate-forming pili (AFP)-encoding genes with the atypical EAEC subgroup. Our study showed that the afp operon was found only in E. coli isolates that carry the aatA gene, but are devoid of the genes encoding the EAEC virulence regulator and the aggregative adherence fimbriae (aatA+/aggR−/AAFs−), thus providing a novel putative marker for increasing the efficiency of atypical EAEC diagnosis.
Data Availability Statement
All datasets generated for this study are included in the article/Supplementary Material.
Ethics Statement
This study was analyzed and approved by the Botucatu Medical School Ethical Committee for human experimentation (CAAE 09994419.6.1001.5411). Written and informed consent from the participants was not required as approved by the aforementioned Ethics Committee.
Author Contributions
RH, LS, and WE conceptualized the study. RD, LS, RH, and WE contributed to the formal analysis. RH and RD were responsible for the funding acquisition. RD, RT, MV, MC-N, LS, TG, WE, and RH carried out the investigation. RD, RT, MV, and MC-N worked on the methodology. RH, RD, and LS helped with the project administration. RH, LS, TG, and WE supervised the study. RH, LS, TG, and WE validated the study. RD and RH wrote the original draft. RD, RT, MV, MC-N, LS, TG, WE, and RH reviewed and edited the manuscript.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001 and CAPES/PROEX 0708/2018.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Cefar Diagnóstica Ltda for providing the antimicrobial drug disks used in this study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2020.00144/full#supplementary-material
References
Andrade, F. B., Abreu, A. G., Nunes, K. O., Gomes, T. A., Piazza, R. M., and Elias, W. P. (2017). Distribution of serine protease autotransporters of Enterobacteriaceae in typical and atypical enteroaggregative Escherichia coli. Infect. Genet. Evol. 50, 83–86. doi: 10.1016/j.meegid.2017.02.018
Andrade, F. B., Gomes, T. A., and Elias, W. P. (2014). A sensitive and specific molecular tool for detection of both typical and atypical enteroaggregative Escherichia coli. J. Microbiol. Methods. 106, 16–18. doi: 10.1016/j.mimet.2014.07.030
Araujo, J. M., Tabarelli, G. F., Aranda, K. R., Fabbricotti, S. H., Fagundes-Neto, U., Mendes, C. M., et al. (2007). Typical enteroaggregative and atypical enteropathogenic types of Escherichia coli are the most prevalent diarrhea-associated pathotypes among Brazilian children. J. Clin. Microbiol. 45, 3396–3399. doi: 10.1128/JCM.00084-07
Aslani, M. M., Alikhani, M. Y., Zavari, A., Yousefi, R., and Zamani, A. R. (2011). Characterization of enteroaggregative Escherichia coli (EAEC) clinical isolates and their antibiotic resistance pattern. Int. J. Infect. Dis. 15, e136–e139. doi: 10.1016/j.ijid.2010.10.002
Baudry, B., Savarino, S. J., Vial, P., Kaper, J. B., and Levine, M. M. (1990). A sensitive and specific DNA probe to identify enteroaggregative Escherichia coli, a recently discovered diarrheal pathogen. J. Infect. Dis. 161, 1249–1251. doi: 10.1093/infdis/161.6.1249
Bernier, C., Gounon, P., and Le Bouguénec, C. (2002). Identification of an aggregative adhesion fimbria (AAF) type III-encoding operon in enteroaggregative Escherichia coli as a sensitive probe for detecting the AAF-encoding operon family. Infect Immun. 70, 4302–4311.doi: 10.1128/iai.70.8.4302–4311.2002
Bhargava, S., Johnson, B. B., Hwang, J., Harris, T. A., George, A. S., Muir, A., et al. (2009). Heat-resistant agglutinin 1 is an accessory enteroaggregative Escherichia coli colonization factor. J. Bacteriol. 191, 4934–4942. doi: 10.1128/JB.01831-08
Boisen, N., Scheutz, F., Rasko, D. A., Redman, J. C., Persson, S., Simon, J., et al. (2012). Genomic characterization of enteroaggregative Escherichia coli from children in Mali. J. Infect. Dis. 205, 431–444. doi: 10.1093/infdis/jir757
Boisen, N., Struve, C., Scheutz, F., Krogfelt, K. A., and Nataro, J. P. (2008). New adhesin of enteroaggregative Escherichia coli related to the Afa/Dr/AAF family. Infect. Immun. 76, 3281–3292.doi: 10.1128/IAI.01646–07
Chattaway, M. A., Day, M., Mtwale, J., White, E., Rogers, J., Day, M., et al. (2017). Clonality, virulence and antimicrobial resistance of enteroaggregative Escherichia coli from Mirzapur, Bangladesh. J. Med. Microbiol. 66, 1429–1435. doi: 10.1099/jmm.0.000594
Chaudhuri, R. R., Sebaihia, M., Hobman, J. L., Webber, M. A., Leyton, D. L., Goldberg, M. D., et al. (2010). Complete genome sequence and comparative metabolic profiling of the prototypical enteroaggregative Escherichia coli strain 042. PLoS ONE 5:e8801. doi: 10.1371/journal.pone.0008801
Clermont, O., Christenson, J. K., Denamur, E., and Gordon, D. M. (2013). The Clermont Escherichia coli phylo-typing method revisited: improvement of specificity and detection of new phylo-groups. Environ. Microbiol. 5, 58–65.doi: 10.1111/1758–2229.12019
Clermont, O., Dixit, O. V. A., Vangchhia, B., Condamine, B., Dion, S., Bridier-Nahmias, A., et al. (2019). Characterization and rapid identification of phylogroup G in Escherichia coli, a lineage with high virulence and antibiotic resistance potential. Environ. Microbiol. 21, 3107–3117. doi: 10.1111/1462-2920.14713
CLSI (2018). Performance Standards for Antimicrobial Susceptibility Testing. CLSI Supplement M100, 28th edn. Wayne, PA: Clinical and Laboratory Standards Institute.
Cravioto, A., Gross, R. J., Scotland, S. M., and Rowe, B. (1979). An adhesive factor found in strains of Escherichia coli belonging to the traditional infantile enteropathogenic serotypes. Curr. Microbiol. 3, 95–99.doi: 10.1007/BF02602439
Croxen, M. A., Law, R. J., Scholz, R., Keeney, K. M., Wlodarska, M., and Finlay, B. B. (2013). Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 26, 822–880.doi: 10.1128/CMR.00022–13
Czeczulin, J. R., Balepur, S., Hicks, S., Phillips, A., Hall, R., Kothary, M. H., et al. (1997). Aggregative adherence fímbria II, a second fimbrial antigen mediating aggregative adherence in enteroaggregative Escherichia coli. Infect. Immun. 65, 4135–4145. doi: 10.1128/IAI.65.10.4135-4145.1997
Czeczulin, J. R., Whittam, T. S., Henderson, I. R., Navarro-Garcia, F., and Nataro, J. P. (1999). Phylogenetic analysis of enteroaggregative and diffusely adherent Escherichia coli. Infect. Immun. 67, 2692–2699. doi: 10.1128/IAI.67.6.2692-2699.1999
Dias, R. C. B., Dos Santos, B. C., Dos Santos, L. F., Vieira, M. A., Yamatogi, R. S., Mondelli, A. L., et al. (2016). Diarrheagenic Escherichia coli pathotypes investigation revealed atypical enteropathogenic E. coli as putative emerging diarrheal agents in children living in Botucatu, São Paulo State, Brazil. APMIS. 124, 299–308. doi: 10.1111/apm.12501
Donnenberg, M. S., Girón, J. A., Nataro, J. P., and Kaper, J. B. (1992). A plasmid-encoded type IV fimbrial gene of enteropathogenic Escherichia coli associated with localized adherence. Mol. Microbiol. 6, 3427–3437. doi: 10.1111/j.1365-2958.1992.tb02210.x
Dudley, E. G., Thomson, N. R., Parkhill, J., Morin, N. P., and Nataro, J. P. (2006). Proteomic and microarray characterization of the AggR regulon identifies a pheU pathogenicity island in enteroaggregative Escherichia coli. Mol. Microbiol. 61, 1267–1282. doi: 10.1111/j.1365–2958.2006.05281.x
Durand, D., Contreras, C. A., Mosquito, S., Ruíz, J., Cleary, T. G., and Ochoa, T. J. (2016). pic gene of enteroaggregative Escherichia coli and its association with diarrhea in Peruvian children. Pathog. Dis. 74:e54. doi: 10.1093/femspd/ftw054
Elias, W. P., Suzart, S., Trabulsi, L. R., Nataro, J. P., and Gomes, T. A. (1999). Distribution of aggA and aafA gene sequences among Escherichia coli isolates with genotypic or phenotypic characteristics, or both, of enteroaggregative E. coli. J. Med. Microbiol. 48, 597–599. doi: 10.1099/00222615–48-6–597
Estrada-Garcia, T., and Navarro-Garcia, F. (2012). Enteroaggregative Escherichia coli pathotype: a genetically heterogeneous emerging foodborne enteropathogen. FEMS Immunol. Med. Microbiol. 66, 281–298. doi: 10.1111/j.1574-695X.2012.01008.x
Ewing, W. H. (1986). Edwards and Ewing's Identification of Enterobacteriaceae, 4th Edn. New York, NY: Elsevier Science Publishing.
França, F. L., Wells, T. J., Browning, D. F., Nogueira, R. T., Sarges, F. S., Pereira, A. C., et al. (2013). Genotypic and phenotypic characterisation of enteroaggregative Escherichia coli from children in Rio de Janeiro, Brazil. PLoS ONE 8:e69971. doi: 10.1371/journal.pone.0069971
Gioppo, N. M., Elias, W. P. Jr., Vidotto, M. C., Linhares, R. E., Saridakis, H. O., Gomes, T. A., et al. (2000). Prevalence of HEp-2 cell-adherent Escherichia coli and characterisation of enteroaggregative E. coli and chain-like adherent E. coli isolated from children with and without diarrhoea, in Londrina, Brazil. FEMS Microbiol. Lett. 190, 293–298. doi: 10.1111/j.1574-6968.2000.tb09301.x
Girón, J. A., Ho, A. S., and Schoolnik, G. K. (1991). An inducible bundle-forming pilus of enteropathogenic Escherichia coli. Science 254, 710–713. doi: 10.1126/science.1683004
Gomes, T. A., Vieira, M. A., Abe, C. M., Rodrigues, D., Griffin, P. M., and Ramos, S. R. (1998). Adherence patterns and adherence-related DNA sequences in Escherichia coli isolates from children with and without diarrhea in São Paulo city, Brazil. J. Clin. Microbiol. 36, 3609–3613. doi: 10.1128/JCM.36.12.3609-3613.1998
Guerrieri, C. G., Monfardini, M. V., Silva, E. A., Bueno de Freitas, L., Schuenck, R. P., and Spano, L. C. (2019). Wide genetic heterogeneity and low antimicrobial resistance of enteroaggregative Escherichia coli isolates from several rural communities. J. Med. Microbiol. 69, 96–103. doi: 10.1099/jmm.0.001120
Harrington, S. M., Dudley, E. G., and Nataro, J. P. (2006). Pathogenesis of enteroaggregative Escherichia coli infection. FEMS Microbiol. Lett. 254, 12–18. doi: 10.1111/j.1574-6968.2005.00005.x
Havt, A., Lima, I. F., Medeiros, P. H., Clementino, M. A., Santos, A. K., Amaral, M. S., et al. (2017). Prevalence and virulence gene profiling of enteroaggregative Escherichia coli in malnourished and nourished Brazilian children. Diagn. Microbiol. Infect. Dis. 89, 98–105. doi: 10.1016/j.diagmicrobio.2017.06.024
Hebbelstrup Jensen, B., Adler Sørensen, C., Hebbelstrup Rye Rasmussen, S., Rejkjær Holm, D., Friis-Møller, A., Engberg, J., et al. (2018). Characterization of diarrheagenic enteroaggregative Escherichia coli in Danish adults-antibiotic treatment does not reduce duration of diarrhea. Front. Cell. Infect. Microbiol. 8:306. doi: 10.3389/fcimb.2018.00306
Hebbelstrup Jensen, B., Olsen, K. E., Struve, C., Krogfelt, K. A., and Petersen, A. M. (2014). Epidemiology and clinical manifestations of enteroaggregative Escherichia coli. Clin. Microbiol. Rev. 27, 614–630. doi: 10.1128/CMR.00112–13
Henderson, I. R., Hicks, S., Navarro-Garcia, F., Elias, W. P., Philips, A. D., and Nataro, J. P. (1999). Involvement of the enteroaggregative Escherichia coli plasmid-encoded toxin in causing human intestinal damage. Infect. Immun. 67, 5338–5344. doi: 10.1128/IAI.67.10.5338-5344.1999
Huang, D. B., Nataro, J. P., DuPont, H. L., Kamat, P. P., Mhatre, A. D., Okhuysen, P. C., et al. (2006). Enteroaggregative Escherichia coli is a cause of acute diarrheal illness: a meta-analysis. Clin. Infect. Dis. 43, 556–563. doi: 10.1086/505869
Ikumapayi, U. N., Boisen, N., Hossain, M. J., Betts, M., Lamin, M., Saha, D., et al. (2017). Identification of subsets of enteroaggregative Escherichia coli associated with diarrheal disease among under 5 years of age children from rural Gambia. Am. J. Trop. Med. Hyg. 97, 997–1004. doi: 10.4269/ajtmh.16-0705
Imuta, N., Ooka, T., Seto, K., Kawahara, R., Koriyama, T., Kojyo, et al. (2016). Phylogenetic analysis of enteroaggregative Escherichia coli (EAEC) isolates from japan reveals emergence of CTX-M-14-producing EAEC O25:H4 clones related to sequence type 131. J. Clin. Microbiol. 54, 2128–2134. doi: 10.1128/JCM.00711-16
Jønsson, R., Liu, B., Struve, C., Yang, Y., Jørgensen, R., Xu, Y., et al. (2017a). Structural and functional studies of Escherichia coli aggregative adherence fimbriae (AAF/V) reveal a deficiency in extracellular matrix binding. Biochim. Biophys. Acta. 1865:304–311. doi: 10.1016/j.bbapap.2016.11.017
Jønsson, R., Struve, C., Boisen, N., Mateiu, R. M., Santiago, A. E., Jenssen, H., et al. (2015). Novel aggregative adherence fimbria variant of enteroaggregative Escherichia coli. Infect. Immun. 83, 1396–1405. doi: 10.1128/IAI.02820-14
Jønsson, R., Struve, C., Boll, E. J., Boisen, N., Joensen, K. G., Sørensen, C. A., et al. (2017b). A novel pAA virulence plasmid encoding toxins and two distinct variants of the fimbriae of enteroaggregative Escherichia coli. Front. Microbiol. 8:263. doi: 10.3389/fmicb.2017.00263
Kaper, J. B., Nataro, J. P., and Mobley, H. L. (2004). Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2, 123–140. doi: 10.1038/nrmicro818
Lang, C., Fruth, A., Holland, G., Laue, M., Mühlen, S., Dersch, P., et al. (2018). Novel type of pilus associated with a Shiga-toxigenic E. coli hybrid pathovar conveys aggregative adherence and bacterial virulence. Emerg. Microbes. Infect. 7:203. doi: 10.1038/s41426-018-0209-8
Lima, A. A. M., Oliveira, D. B., Quetz, J. S., Havt, A., Prata, M. M. G., Lima, I. F. N., et al. (2019a). Etiology and severity of diarrheal diseases in infants at the semiarid region of Brazil: a case-control study. PLoS Negl. Trop. Dis. 13:e0007154. doi: 10.1371/journal.pntd.0007154
Lima, F. M., de Paulo Daurelio, F., Mucci, E. R., Ahagon, C. M., Dos Santos Carmo, A. M., Eterovic, A., et al. (2019b). Epidemiology and genetic screening of diarrheagenic Escherichia coli among symptomatic and asymptomatic children. J. Med. Microbiol. 68, 1033–1041. doi: 10.1099/jmm.0.001020
Lima, I. F., Boisen, N., Quetz Jda, S., Havt, A., de Carvalho, E. B., Soares, A. M., et al. (2013). Prevalence of enteroaggregative Escherichia coli and its virulence-related genes in a case-control study among children from north-eastern Brazil. J. Med. Microbiol. 62, 683–693. doi: 10.1099/jmm.0.054262–0
Lozer, D. M., Souza, T. B., Monfardini, M. V., Vicentini, F., Kitagawa, S. S., Scaletsky, I. C. A., et al. (2013). Genotypic and phenotypic analysis of diarrheagenic Escherichia coli strains isolated from Brazilian children living in low socioeconomic level communities. BMC Infect. Dis. 13:418. doi: 10.1186/1471-2334-13-418
Magiorakos, A. P., Srinivasan, A., Carey, R. B., Carmeli, Y., Falagas, M. E., Giske, C. G., et al. (2012). Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 18, 268–281. doi: 10.1111/j.1469-0691.2011.03570.x
Monteiro-Neto, V., Bando, S. Y., Moreira-Filho, C. A., and Girón, J. A. (2003). Characterization of an outer membrane protein associated with haemagglutination and adhesive properties of enteroaggregative Escherichia coli O111:H12. Cell. Microbiol. 5, 533–547. doi: 10.1046/j.1462–5822.2003.00299.x
Morin, N., Santiago, A. E., Ernst, R. K., Guillot, S. J., and Nataro, J. P. (2013). Characterization of the AggR regulon in enteroaggregative Escherichia coli. Infect. Immun. 81, 122–132. doi: 10.1128/IAI.00676–12
Nataro, J. P., Deng, Y., Maneval, D. R., German, A. L., Martin, W. C., and Levine, M. M. (1992). Aggregative adherence fimbriae I of enteroaggregative Escherichia coli mediate adherence to HEp-2 cells and hemagglutination of human erythrocytes. Infect. Immun. 60, 2297–2304. doi: 10.1128/IAI.60.6.2297-2304.1992
Nataro, J. P., and Kaper, J. B. (1998). Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 11, 142–201. doi: 10.1128/CMR.11.1.142
Nataro, J. P., Kaper, J. B., Robins-Browne, R., Prado, V., Vial, P., and Levine, M. M. (1987). Patterns of adherence of diarrheagenic Escherichia coli to HEp-2 cells. Pediatr. Infect. Dis. J. 6, 829–831. doi: 10.1097/00006454-198709000-00008
Nishi, J., Sheikh, J., Mizuguchi, K., Luisi, B., Burland, V., Boutin, A., et al. (2003). The export of coat protein from enteroaggregative Escherichia coli by a specific ATP-binding cassette transporter system. J. Biol. Chem. 278, 45680–45689. doi: 10.1074/jbc.M306413200
Ori, E. L., Takagi, E. H., Andrade, T. S., Miguel, B. T., Cergole-Novella, M. C., Guth, B. E. C., et al. (2019). Diarrhoeagenic Escherichia coli and Escherichia albertii in Brazil: pathotypes and serotypes over a 6-year period of surveillance. Epidemiol. Infect. 147, 1–9. doi: 10.1017/S0950268818002595
Rodrigues, J., Scaletsky, I. C., Campos, L. C., Gomes, T. A., Whittam, T. S., and Trabulsi, L. R. (1996). Clonal structure and virulence factors in strains of Escherichia coli of the classic serogroup O55. Infect. Immun. 64, 2680–2686. doi: 10.1128/IAI.64.7.2680-2686.1996
Santiago, A. E., Yan, M. B., Hazen, T. H., Sauder, B., Meza-Segura, M., Rasko, D. A., et al. (2017). The AraC Negative Regulator family modulates the activity of histone-like proteins in pathogenic bacteria. PLoS Pathog. 13:e1006545. doi: 10.1371/journal.ppat.1006545
Scaletsky, I. C., Fabbricotti, S. H., Aranda, K. R., Morais, M. B., and Fagundes-Neto, U. (2002). Comparison of DNA hybridization and PCR assays for detection of putative pathogenic enteroadherent Escherichia coli. J. Clin. Microbiol. 40, 1254–1258. doi: 10.1128/JCM.40.4.1254-1258.2002
Scaletsky, I. C., Silva, M. L., and Trabulsi, L. R. (1984). Distinctive patterns of adherence of enteropathogenic Escherichia coli to HeLa cells. Infect. Immun. 45, 534–536. doi: 10.1128/IAI.45.2.534-536.1984
Sheikh, J., Czeczulin, J. R., Harrington, S., Hicks, S., Henderson, I. R., Le Bouguénec, C., et al. (2002). A novel dispersin protein in enteroaggregative Escherichia coli. J. Clin. Invest. 110, 1329–1337. doi: 10.1172/JCI16172
Sheikh, J., Dudley, E. G., Sui, B., Tamboura, B., Suleman, A., and Nataro, J. P. (2006). EilA, a HilA-like regulator in enteroaggregative Escherichia coli. Mol. Microbiol. 61, 338–350. doi: 10.1111/j.1365-2958.2006.05234.x
Spano, L. C., Cunha, K. F., Monfardini, M. V., Fonseca, R. C. B., and Scaletsky, I. C. A. (2017). High prevalence of diarrheagenic Escherichia coli carrying toxin-encoding genes isolated from children and adults in southeastern Brazil. BMC Infect. Dis. 17:773. doi: 10.1186/s12879-017-2872-0
Taborda, R. L. M., Silva, L. A. D., Orlandi, P. P., Batista, F. S., Rodrigues, R. S., and Matos, N. B. (2018). Characterization of enteroaggregative Escherichia coli among diarrheal children in western brazilian amazon. Arq. Gastroenterol. 55, 390–396. doi: 10.1590/s0004-2803.201800000-84
Tobe, T., Hayashi, T., Han, C. G., Schoolnik, G. K., Ohtsubo, E., and Sasakawa, C. (1999). Complete DNA sequence and structural analysis of the enteropathogenic Escherichia coli adherence factor plasmid. Infect. Immun. 67, 5455–5462. doi: 10.1128/IAI.67.10.5455-5462.1999
Keywords: EAEC, virulence factors, diarrhea, diagnosis, antimicrobial resistance
Citation: Dias RCB, Tanabe RHS, Vieira MA, Cergole-Novella MC, dos Santos LF, Gomes TAT, Elias WP and Hernandes RT (2020) Analysis of the Virulence Profile and Phenotypic Features of Typical and Atypical Enteroaggregative Escherichia coli (EAEC) Isolated From Diarrheal Patients in Brazil. Front. Cell. Infect. Microbiol. 10:144. doi: 10.3389/fcimb.2020.00144
Received: 06 February 2020; Accepted: 19 March 2020;
Published: 22 April 2020.
Edited by:
Guoquan Zhang, The University of Texas at San Antonio, United StatesReviewed by:
George P. Munson, University of Miami, United StatesJorge Blanco, University of Santiago de Compostela, Spain
Copyright © 2020 Dias, Tanabe, Vieira, Cergole-Novella, dos Santos, Gomes, Elias and Hernandes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rodrigo T. Hernandes, rt.hernandes@unesp.br