
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol. , 21 January 2020
Sec. Parasite and Host
Volume 9 - 2019 | https://doi.org/10.3389/fcimb.2019.00477
This article is part of the Research Topic The Role of Saliva in Arthropod-Host-Pathogen Relationships View all 15 articles
 Poliana Fernanda Giachetto1
Poliana Fernanda Giachetto1 Rodrigo Casquero Cunha2
Rodrigo Casquero Cunha2 Antônio Nhani Jr.1Marcos Valerio Garcia3
Antônio Nhani Jr.1Marcos Valerio Garcia3 Jesus Aparecido Ferro4
Jesus Aparecido Ferro4 Renato Andreotti3*
Renato Andreotti3*The success of cattle tick fixation largely depends on the secretion of substances that alter the immune response of the host. The majority of these substances are expressed by the parasite salivary gland and secreted in tick saliva. It is known that hosts can mount immune responses against ticks and bovine European breeds, and bovine industrial crossbreeds are more susceptible to infestations than are Bos indicus cattle. To identify candidates for the development of novel control strategies for the cattle tick Rhipicephalus (Boophilus) microplus, a salivary gland transcriptome analysis of engorged females fed on susceptible or resistant hosts was performed. Using RNA-Seq, transcriptomes were de novo assembled and produced a total of 235,451 contigs with 93.3% transcriptome completeness. Differential expression analysis identified 137 sequences as differentially expressed genes (DEGs) between ticks raised on tick-susceptible or tick-resistant cattle. DEGs predicted to be secreted proteins include innexins, which are transmembrane proteins that form gap junction channels; the transporters Na+/dicarboxylate, Na+/tricarboxylate, and phosphate transporter and a putative monocarboxylate transporter; a phosphoinositol 4-phosphate adaptor protein; a cysteine-rich protein containing a trypsin inhibitor-like (TIL) domain; a putative defense protein 3 containing a reeler domain; and an F-actin-uncapping protein LRRC16A with a CARMIL_C domain; these genes were upregulated in ticks fed on tick-susceptible cattle. DEGs predicted to be non-secreted proteins included a small heat shock protein and the negative elongation factor B-like, both acting in a coordinated manner to increase HSP transcript levels in the salivary glands of the ticks fed on tick-susceptible cattle; the 26S protease regulatory subunit 6B and another chaperone with similarity to calnexin, also upregulated in ticks fed on tick-susceptible cattle; an EF-hand calcium binding protein and a serine carboxypeptidase (SCP), both involved in the blood coagulation cascade and upregulated in ticks fed on tick-susceptible cattle; and two ribosomal proteins, the 60S acidic ribosomal protein P2 and the 60S ribosomal protein L19. These results help to characterize cattle tick salivary gland gene expression in tick-susceptible and tick-resistant hosts and suggest new putative targets for the control of tick infestations, as those genes involved in the mechanism of stress response during blood feeding.
The cattle tick R. (B.) microplus limits the development of the cattle industry worldwide, causing production losses estimated at US $3.24 billion annually in Brazil alone (Grisi et al., 2014). The losses caused by ticks are caused primarily by their feeding in the host and by pathogens transmitted via saliva thereafter. To feed, the tick must attach to the skin of the cattle, introducing their hypostome. The success of the fixation of the tick depends on the secretion of cement substances and anticoagulants, which alter the immune response in the place of the bite but can also cause systemic effects (Mans and Neitz, 2004). In addition, the success of pathogen transmission depends on some tick molecules associated with this event (Ramamoorthi et al., 2005; Hovius et al., 2008). The majority of these substances are expressed by the salivary gland and may be secreted in the saliva.
The tick saliva contains a rich variety of pharmacologically bioactive molecules that support blood feeding. During coevolution, blood sucking ticks have adapted mechanisms to evade host detection and prevent blood coagulation by synthesizing an extensive array of molecules with anesthetic, immunosuppressive, vasodilatory, profibrinolytic, and anticoagulant properties (Mans and Neitz, 2004). Blood feeding triggers a heat shock response by arthropods, as demonstrated by the increased production of heat shock proteins in response to the increase in temperature and other stresses observed during blood meal by ticks, which has been considered a stressful event in multiple forms (Shahein et al., 2010; Benoit et al., 2011).
Gene transcripts, such as glutathione S-transferase and gamma-glutamyl transferase, can be found in salivary glands because they have physiological functions; one of these genes plays a central role in the detoxication of xenobiotic compounds (de Lima et al., 2002), such as insecticides (Nandi et al., 2015; Hernandez et al., 2018), and another of which is involved in the cross-cell membrane trafficking of amino acids and peptides and in glutathione metabolism, respectively (Mulenga and Erikson, 2011).
Many other transcripts that can code for non-secreted or secreted proteins with different physiological functions may be present in tick salivary glands. Examples of predicted non-secreted proteins are the following: calnexin, which plays a role in the quality control and assembly of proteins and glycoproteins in the endoplasmic reticulum (Williams, 2006); longistatin, which modulates biochemical reactions within the cell as the inflammatory response and has a role in anticoagulant action (Anisuzzaman et al., 2012); serine carboxypeptidase, implied to be involved in degrading hemoglobin to peptides and regulating the interaction with the host; β-N-acetyl hexosaminidases, which participates in the turnover of the chitin exoskeleton (Hogenkamp et al., 2008); leucine aminopeptidase, which belongs to a diverse group of the M17 family of Zn-metalloproteases (Maggioli et al., 2018), playing important roles in the host immune response, tick-tissue development, and pathogen transmission (Ali et al., 2015); ribosomal proteins, playing essential roles in cell growth and proliferation (Trainor and Merrill, 2014); phosphorylase kinase, a holoenzyme that activates glycogen phosphorylase (Brushia and Walsh, 1999); E3 ligase, promoting cullin neddylation, required for the regulation of NF-κB, which is crucial in immune response and apoptotic pathways (Cajee et al., 2012); and mucins, providing lubrication and protection to the epithelium (Hollingsworth and Swanson, 2004).
Among the salivary gland transcripts predicted to code for secreted proteins, the following genes may be observed: innexins, transmembrane proteins (Richards et al., 2015) that form gap junction channels and hemichannels in invertebrates, including arthropods (Güiza et al., 2018); Na+/dicarboxylate, Na+/tricarboxylate and phosphate transporter, which may play a role in osmoregulation with ion transport function (Hui et al., 2014); the phosphoinositol 4-phosphate adaptor protein, a component of a molecule complex that recruits proteins to the cell membrane (Choudhury et al., 2005), a transduction pathway in tick salivary glands (McSwain et al., 1989); monocarboxylate transporters that catalyze rapid transport of many monocarboxylates (Lew-Tabor et al., 2011), cysteine-rich proteins containing trypsin inhibitor-like (TIL) domain, belonging to the family that comprises chymotrypsin, elastase and trypsin inhibitors (Sasaki et al., 2008), which has been found ubiquitously in blood-feeding insect and tick sialomes (Karim and Adamson, 2012; Maruyama et al., 2017); BmSI-7, a trypsin inhibitor-like cysteine-rich domain family, which is involved in the inflammatory response and in injury caused by tick fixation on the bovines (Sasaki et al., 2008); defense proteins, such as defense protein 3 (Zhao et al., 2019), and ixodidin, a antimicrobial peptide (Fogaça et al., 2006), which are probably involved in the immune response in the tick; PIXR, a protein with a reeler domain, favoring the colonization of Borrelia burgdorferi in Ixodes scapularis gut (Narasimhan et al., 2017); and proteins involved in actin metabolism, such as the F-actin-uncapping protein LRRC16A isoform X2.
All of these compounds may play a role interacting on the interface of the tick-host relation and are crucial to the success of tick fixation and/or pathogen transmission. On this basis, knowledge of the transcripts that have been upregulated or downregulated in both tick-susceptible and tick-resistant cattle could guide the development of more consistent immunogens against ticks at the time when the use of vaccines has presented results with varied effects, and the type of antigen used interferes with the efficacy of the vaccine (Csordas et al., 2018). In the absence of a vaccine with these characteristics, only chemical control remains for cattle producers seeking to prevent tick infestations. In addition, the use of acaricides at higher concentrations and in association to overcome the resistance of the tick to the acaricides indicates a global crisis in tick control (Higa et al., 2019), and vaccine development is a promising alternative.
It has been known for many years that hosts can mount immune responses against ticks, and non-natural hosts can mount highly effective anti-tick immune responses, preventing successful tick fixation and halting the tick life cycle (Trager, 1939). In this way, bovine European breeds (Bos taurus) and their industrial crossbreeds are more susceptible to tick infestations than B. indicus. This resistance is associated with the immune systems of the hosts, since in the first infestation, the number of ticks completing the cycle is similar in all races (Hewetson, 1972; Mattioli et al., 1993; Ghosh et al., 1999), and with the absence of transcripts of genes encoding enzymes producing volatile compounds, which may render the host less attractive to larvae (Franzin et al., 2017). Franzin et al. (2017) concluded that resistant hosts expose ticks to an earlier inflammatory response, which in ticks is associated with significantly lower expression of genes encoding salivary proteins that suppress host immunity, inflammation and coagulation. It means, at the same time, different levels of host immunity may affect the composition of tick saliva, contributing to these outcomes. The demonstration that immunosuppressive drugs eliminate this resistance reaffirmed the immune nature of this response (Bergman et al., 2000).
In this context, we aimed to differentiate the profile of salivary gland transcripts in ticks feeding on tick-resistant and tick-susceptible bovines to identify transcripts of genes encoding proteins with the potential to be used as immunogens for tick control.
All experimental procedures were approved by Embrapa Beef Cattle's Ethics Committee on Animal Use according to Protocol 008/2014 and according to the requirements of the National Council for the Control of Animal Experimentation.
The trial for sample collection was conducted in Campo Grande, Mato Grosso do Sul, Brazil (20° 27′ S and 54° 37′ W, altitude of 530 m), at Embrapa Beef Cattle. The climate of this region is classified as rainy tropical savanna, characterized by irregular annual rainfall distribution, a well-defined dry period during the colder months, and a rainy period during the summer months.
Two weaned male cattle aged ~8 months from two bovine genotypes were used: a tick-susceptible animal, Holstein (Bos taurus taurus), and a tick-resistant animal, crossbreed, resulting from the crossing of the previous susceptible breed with a tick-resistant breed, Nelore (Bos taurus indicus). The animals were kept in a Brachiaria decumbens pasture for a 4 months period for disinfestation and immunological memory loss, and they were later taken to individual stalls, where they received feed of sorghum silage (17 kg/animal/day), concentrate (1 kg/animal/day), and water ad libitum throughout the experimental period.
Ticks used in the experiment for artificial infestation were from a colony of R. (B.) microplus reared in the Laboratory of Tick Biology of Embrapa Beef Cattle. Engorged females from a stabled bovine were collected (21 days) placed in Petri dishes and incubated in a biological oxygen demand chamber (BOD) at a temperature of ~28°C and humidity of ~80%. After egg laying, eggs were collected and separated into 500-mg samples containing an equivalent of 10,000 larvae. Samples were placed in 10 ml syringes with the tips removed, sealed with cotton, and then placed again on BOD for egg incubation until larval hatching. Syringes containing non-fed larvae were used for artificial infestation of cattle. An infestation with 10,000 active larvae (12 days after hatching) was performed on the dorsum in each of the experimental animals—the tick-susceptible and tick-resistant cattle, which remained immobilized for a period of 3 h to avoid self-cleaning.
Semi-engorged adult salivary gland samples were dissected on ice and processed immediately. Three salivary gland samples were obtained for each of the bovine genotypes. In a total of six samples, each sample contained salivary glands collected from ~10 ticks, which were used for RNA isolation following the TRIzol® reagent protocol (Life Technologies, Carlsbad, CA). The purity and amount of total RNA were assessed by electrophoresis in a 1.2% agarose gel stained with ethidium bromide and by spectrophotometry at 260 and 280 nm in a Nanodrop ND-1000 spectrophotometer (Thermo Fisher Scientific Inc., MA, USA). Total RNA quality and integrity were analyzed using an Agilent 2100 Bioanalyzer® (Agilent Technologies, Palo Alto, USA). RNA was stored at −70°C until further analysis.
Library preparation and next generation sequencing using the RNA-Seq technique were conducted at the Animal Biotechnology Laboratory of Luiz de Queiroz College of Agriculture, University of São Paulo (ESALQ-USP, Piracicaba, SP), following the stranded TruSeq RNA Sample Prep Kit protocol (Illumina, Inc., San Diego, CA, USA). The cDNA libraries were sequenced in the Illumina HiSeq 2500 System (Illumina, Inc., San Diego, CA, USA), generating 2 × 150 bp paired-end reads, according to the standard manufacturer protocol.
Data obtained by HiSeq 2500 sequencing were analyzed using the Real Time Analysis (Illumina) software, which makes the calls from the sequencing images, converting them into a FASTQ format. Evaluation of the sequencing reads was performed using FastQC v0.11.4 (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Read sequences were subjected to adapter trimming and quality filtering using Trimmomatic (v0.35) (Bolger et al., 2014) with default parameters, except for a head crop of 15 bases and a read minimal length of 30 bases. Reads were also checked for foreign RNA contamination. A similarity search against the NR database (v04/18) using either blastx (v2.6.0) or DIAMOND (v0.9.17) (Buchfink et al., 2015) was performed to identify and filter hits to mammal and bacterial sequences above 50% identity and e-value ≤ 1e-20.
Trinity software (v2.6.6) (Grabherr et al., 2011) was used to de novo assembly of a reference transcriptome using all filtered samples, as described in https://github.com/trinityrnaseq/trinityrnaseq/wiki. Inchworm kmer coverage was set to 3, and contig minimal length was set to 300 bases. Assembly quality was accessed through mapping back each library to the R. (B.) microplus salivary gland reference transcriptome using Bowtie2 (v2.3.4) (Langmead and Salzberg, 2012) and Trinity scripts. Mapped contig sequences were recovered for each genotype to compose the transcriptomes.
The completeness of the R. (B.) microplus salivary gland transcriptome was evaluated using BUSCO (Benchmarking Universal Single-Copy Orthologs) by comparing the transcriptome against a set of highly conserved single-copy orthologs of the known ancestral Arthropoda proteins (arthropoda_odb9, creation date: 2017-02-07, number of species: 60, number of BUSCOs: 1066) (Simão et al., 2015).
Differentially expressed genes (DEGs) between ticks fed on susceptible cattle and ticks fed on resistant cattle were identified by Trinity RSEM/edgeR differential expression analysis with FDR (False Discovery Rate) <0.05 and Fold Change ≥2. A preliminary similarity search, as described previously, was also performed in the DEG set of transcripts.
TransDecoder (v 5.0.0) (Haas et al., 2013) was used to obtain peptide ORFs from differentially expressed genes. Potential signal peptides and transmembrane domains were predicted from ORFs to identify secreted proteins according to the Min (2010) animal pipeline, which consists of filtering sequences that pass through Phobius (v 1.01), WolfPsort (v 0.02) TMHMM (v 2.0c) TargetP, and PS_SCAN tools.
To further investigate the biological functions of the DEGs, they were annotated to the Gene Ontology (GO) database for biological process (BP), molecular function (MF), and cellular component (CC) and to KEGG (Kyoto Encyclopedia of Genes and Genomes) for biological pathways using Blast2GO PRO (https://www.blast2go.com/). DEGs from the most represented GO terms and KEGG pathways were discussed further.
Sialotranscriptomes from R. (B.) microplus fed on tick-susceptible (Holstein) and tick-resistant (Holstein × Nelore crossbreed) cattle were generated using RNA-Seq technology and de novo assembly. A total of 74,639,552 reads were obtained for ticks fed on Holstein cattle, and 63,013,658 reads were obtained for ticks fed on crossbreed cattle. Reads post-processing with Trimommatic resulted in a 99.9% recovery for the Holstein and crossbreed reads dataset, of which 92.53 and 92.79%, respectively, were realigned to the assembly with bowtie2. Trinity produced a total of 235,451 contigs with a mean length of 954 nt and a N50 of 1,624 nt. Completeness of the assembly, as evaluated using BUSCO, revealed that 93.3% of conserved genes across Arthropoda were present (994 out of 1,066). Complete and single-copy genes found (C) were 24.4%, complete and duplicated genes found (D) were 68.9%, fragmented genes found (F) were 5.0%, and genes missing (M) were 1.7%. To identify the function of the transcriptome-predicted genes, all 235,451 contigs were analyzed using Blast2GO PRO software, returning a total of 71,757 annotated transcripts with a sequence identity ≥50% and e-value ≤1e−10 (Table 1, Supplementary Material). Gene Ontology and KEGG categorizations of the annotated transcriptome are shown in Figure 1.
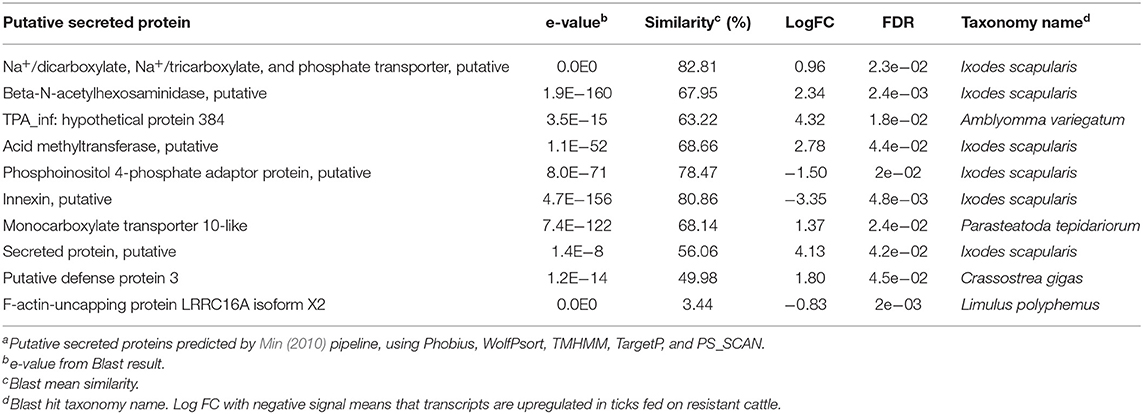
Table 1. Putative secreted proteinsa transcribed in R. (B.) microplus salivary glands.
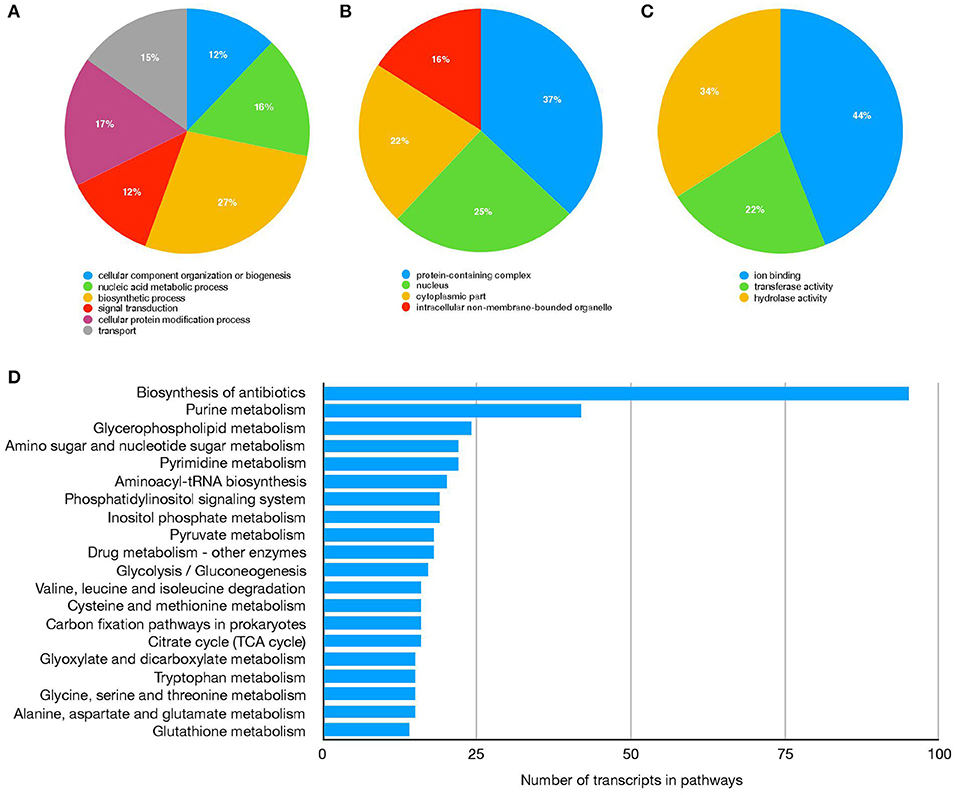
Figure 1. R. (B.) microplus salivar gland transcriptome functional annotation. Transcripts distribution based on GO categories Biological process (A), Cellular component (B), and Molecular function (C). Top 20 processes in KEGG pathways (D).
TransDecoder translation resulted in a total of 1,815 predicted ORFs, which were used for secreted protein identification. According to the pipeline proposed by Min (2010), a total of 20 ORFs were predicted as putative secreted proteins from the salivary glands (Table 1), corresponding to 15 candidate genes.
The differential expression analysis identified 137 sequences as DEGs, corresponding to 126 candidate genes (Figure 1, Table 2, Supplementary Material). Gene Ontology analysis showed that the molecular functions most represented among DEGs upregulated in salivary glands from R. (B.) microplus fed on tick-susceptible and tick-resistant cattle were ion binding and hydrolase activity, while the most represented biological processes were the nitrogen compound metabolic process and the cellular metabolic process (Figure 2 and Table 2).
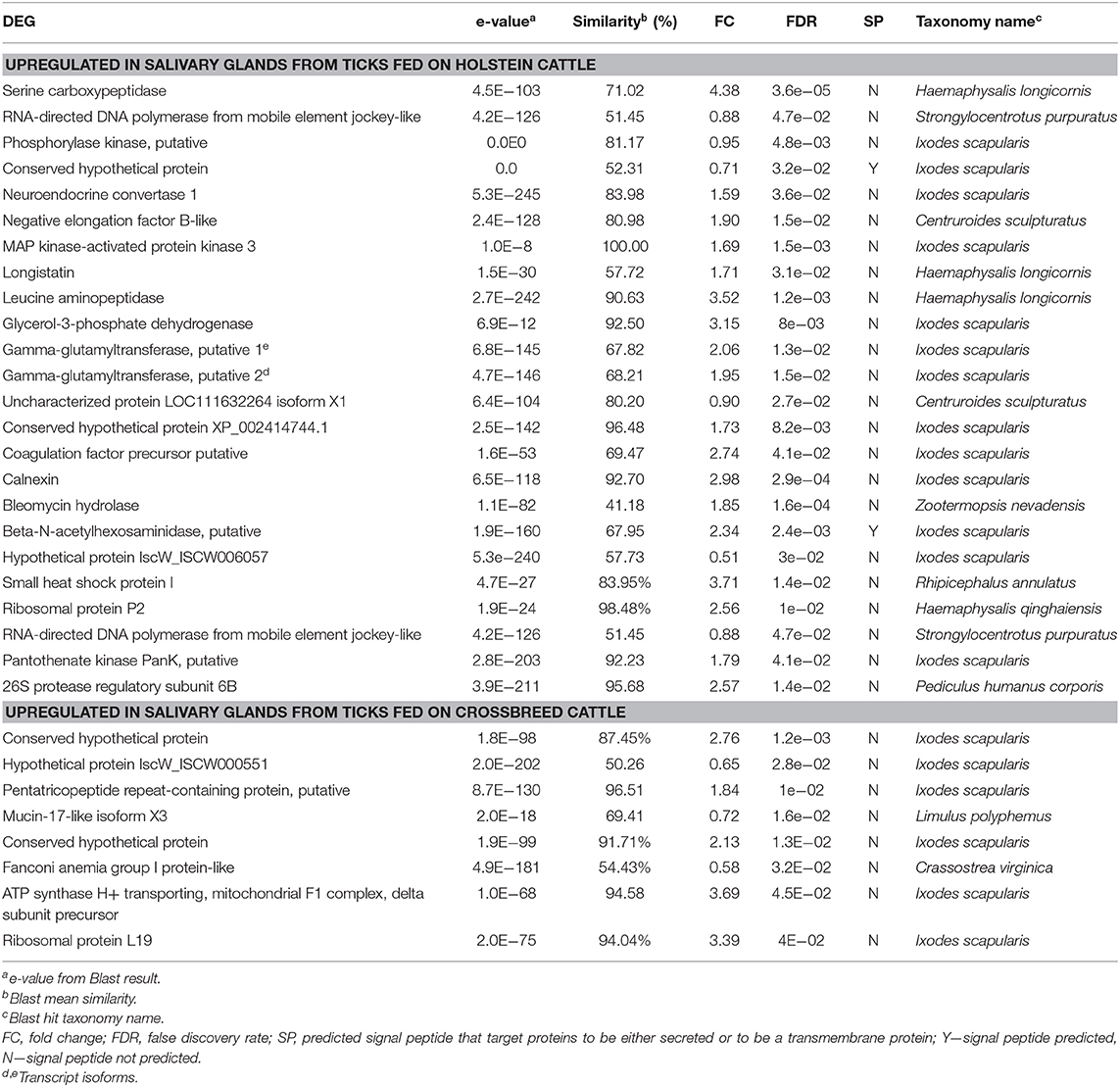
Table 2. Differentially expressed genes (DEG) upregulated in salivary glands from ticks fed on susceptible-tick cattle (Holstein) and ticks fed on resistant-tick cattle (Holstein × Nelore crossbreed) from most represented molecular functions and biological process GO terms.
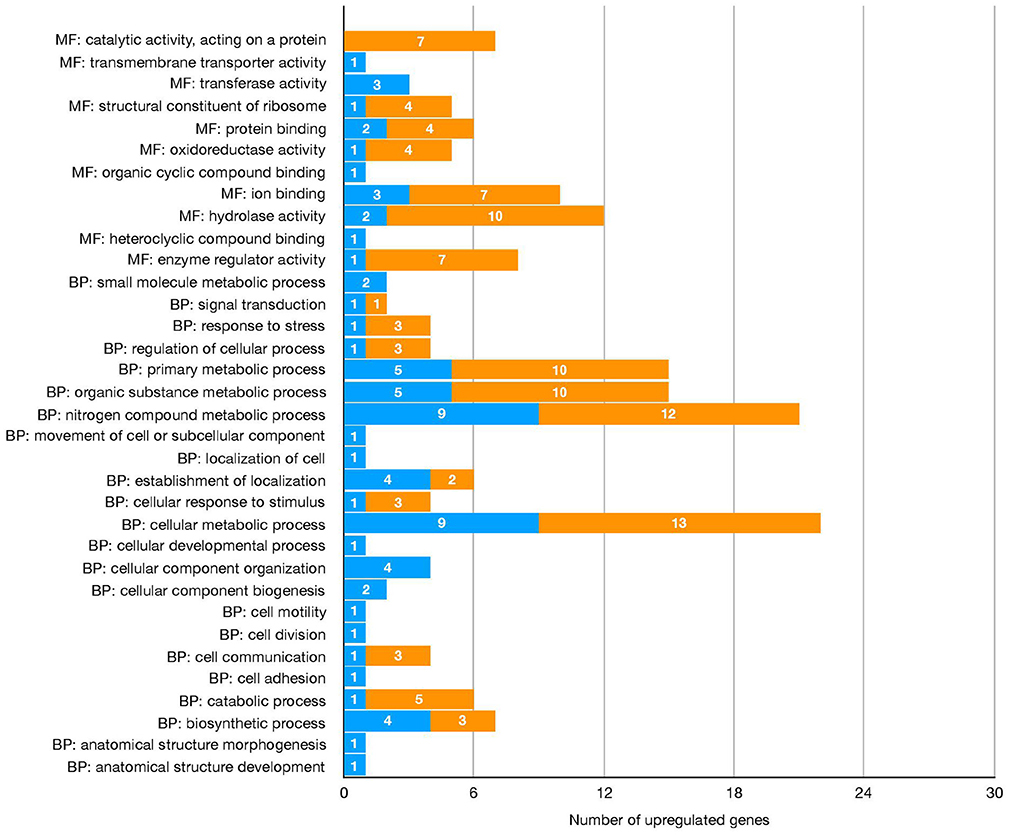
Figure 2. Number of the differentially expressed genes upregulated in salivary glands of R. (B.) microplus fed on Holstein (susceptible) and crossbreed (resistant) cattle, based on GO categories Molecular function (MF) and Biological process (BP).
Blood feeding in arthropods is a stressful event in multiple forms and physiological targets. Energetic, thermal, osmotic and oxidative stresses are among the consequences of the rapid ingestion of large amounts of warm blood (Pereira et al., 2017). The large volumes of blood that arrives quickly to the midgut may increase the temperature of blood-feeding arthropods by up to 15°C in <1 min, and the thermal stress generated by the blood meal can trigger a heat shock response by arthropods, as demonstrated by the increased production of heat shock proteins (Benoit et al., 2011).
A transcript upregulated in ticks fed on tick-susceptible cattle showed similarity with the sequence of a small heat shock protein I gene cloned from salivary glands of Rhipicephalus annulatus—the RasHSPI (Hussein et al., 2014). The authors verified a strong immunogenic effect of the RasHSPI recombinant protein in rabbits, suggesting that it could be used as a potential protective antigen (Hussein et al., 2014). Previously, another sHSP also cloned from R. annulatus salivary glands, a RasHSPII, showed a role as a molecular chaperone, conferring protection at least or specifically to the tick salivary glands against the increase in temperature and other stresses observed during blood feeding by ticks (Shahein et al., 2010). Guilfoile and Packila (2004) verified an increase in the expression of a gene with similarity to a HSP70 in I. scapularis females and males throughout blood feeding and suggested that protein encoded by the gene may be required to stabilize tick proteins during ingestion of warm blood from the host or even when the parasite is in contact with the hot skin of the host. An increase in the expression of a small heat shock protein, HSP16, was also observed during the engorgement of I. scapularis females (Xu et al., 2005). According to Benoit et al. (2019), suppression of HSPs through RNA interference (RNAi) also leads to impaired digestion and subsequent egg production, indicating that prevention or repair of thermal-associated damage is critical to allow blood feeding arthropods to maximize egg production from a blood meal.
Recently, the role of an arthropod HSP70-like molecule in fibrinogenolysis during blood meal has been reported (Vora et al., 2017), as well as the protective effect of cellular protein integrity observed with the overexpression of genes from the heat shock protein family. In our results, in addition to an increase in heat shock gene expression observed in ticks fed on tick-susceptible cattle, downregulation of a transcript similar to the negative elongation factor B-like (NELF) was observed. Ghosh et al. (2011) found that NELF depletion in Drosophila, mediated by RNA interference, resulted in a delay in the dissociation of heat shock factors (HSF), the transcription factor that upregulates genes encoding heat shock proteins of the HSP genes during heat shock recovery. Thus, the observed increase in HSP transcript levels in the salivary gland of the ticks fed on tick-susceptible cattle may be related to the delay in the dissociation of the HSF from the HSP genes due to the downregulation of the NELF. This coordinated action may contribute to the success of the parasite in its interaction with the host.
Two isoforms of a transcript with similarity to an I. scapularis gamma-glutamyl transferase (GGT) were upregulated in ticks fed tick-susceptible cattle. GGT is an enzyme present in cell membranes and is involved in cross-cell-membrane trafficking of amino acids and peptides and in glutathione metabolism (Mulenga and Erikson, 2011). Glutathione S-transferases (GST) are proteins that belong to a multigene family and play a central role in the detoxification of xenobiotic compounds (de Lima et al., 2002), with increased expression levels being correlated with insecticide resistance in various organisms, including ticks (Nandi et al., 2015; Hernandez et al., 2018). GGT plays key roles in intracellular reduced glutathione (GSH) homeostasis by breaking down the extracellular molecule and providing cysteine for intracellular de novo synthesis of GSH (Accaoui et al., 2000; Zhang et al., 2005). Glutathione biosynthesis by GGT is important to maintain intracellular reduced glutathione and the cellular redox state. Additionally, during oxidative stress in rodents, GGT gene expression was increased, which is believed to constitute an adaptation to stress (Zhang et al., 2005). The authors also related that increased expression of GGT during oxidative stress facilitates GSH turnover, de novo GSH synthesis, and metabolism and detoxification of GSH conjugates, increasing cell resistance to subsequent stress. In Haemaphysalis longicornis, the analysis of two GSTs using real-time PCR showed that gene expression increases continuously in salivary glands and other organs of the tick as blood-feeding progresses until female engorgement, demonstrating the possible role of GSTs in coping with oxidative stress caused by blood-feeding (Hernandez et al., 2018).
The 26S proteasome, the heart of the ubiquitin-proteasome system, is a catalytic machine that cleaves most intracellular ubiquitinated proteins, maintaining cellular homeostasis by means of the recognition, unfolding, translocating and cleavage of protein substrates (Wang and Maldonado, 2006; Gallastegui and Groll, 2010). This proteasome is involved in the regulation of many crucial processes in the cell, such as the cell cycle, apoptosis, signal transduction, protein processing and immune and stress responses (Coux et al., 1996). In our study, the 26S protease regulatory subunit 6B was upregulated in salivary glands from ticks fed tick-susceptible cattle. An increase in the abundance of proteasome proteins was verified in Trypanosoma cruzy exposed to the antiparasitic benznidazole (Andrade et al., 2008). In the small brown planthopper Laodelphgax striatellus, the vector of the rice stripe virus, an increase in the accumulation of the virus was observed after disrupting the 26S proteasome, indicating that the small brown planthopper 26S proteasome plays a role in defense against rice stripe virus infection by regulating virus accumulation (Xu et al., 2015). In vertebrates, another function associated with proteasome proteolytic activity, first suggested by Yang et al. (1992), is that the proteasome is also a proteolytic generator of antigenic peptides that can be presented by class I MHC molecules. A 26S proteasome non-ATPase regulatory subunit protein was characterized as an immunogenic enzyme in 24 h fed I. scapularis saliva (Lewis et al., 2015), and in H. longicornis, the increased expression of the 26S proteasome regulatory subunit protein indicated an increase in expression of the ubiquitin-proteasome system during salivary gland degeneration in engorged females, certainly as a consequence of the programmed cell death that salivary glands undergo after tick engorgement (Wang et al., 2019).
Another chaperone, a transcript with similarity to calnexin (CNX), was upregulated in ticks fed tick-susceptible cattle. Calnexin is a chaperone involved in the quality control of proteins, ensuring correct folding and assembly of many glycoproteins in the endoplasmic reticulum (Williams, 2006). The endoplasmic reticulum is a specialized organelle that plays essential and central functions in the cell, including protein folding assisted by a battery of molecular chaperones. Situations that reduce the protein folding capacity of the endoplasmic reticulum, which results in the accumulation and aggregation of unfolded proteins, cause a condition known as endoplasmic reticulum stress (Guérin et al., 2008). In Schyzosaccharomyces pombe, calnexin overexpression is involved in the induction of apoptosis triggered by endoplasmic reticulum stress (Guérin et al., 2008). In the midgut of the argasid tick Ornithodoros moubata female, a calnexin protein has been identified and determined to be involved with the stress responses associated with blood digestion (Oleaga et al., 2017). A calnexin transcript was upregulated in the Ixodes ricinus-derived cell line IRE/CTVM19 at day 2 post-infection with the flavivirus tick-borne encephalitis virus (TBEV), which may have an important role in the response of tick cells to virus infection (Weisheit et al., 2015). In R. (B.) microplus, calnexin was predicted as a transmembrane protein and has been suggested as a target for tick control strategies because it showed a higher VaxiJen score (0.91) than that predicted for Bm86 (0.77) in a study from Richards et al. (2015). Calnexin has also been suggested as a promising vaccine candidate against multiple fungal pathogens because it induces fungal antigen-specific CD4+ T cell expansion and resistance to lethal challenge in mice with multiple fungal pathogens (Wüthrich et al., 2015).
The tick saliva contains a rich variety of pharmacologically bioactive molecules that support blood feeding. During coevolution, blood sucking ticks have adapted mechanisms to evade host detection and prevent blood coagulation by synthesizing an extensive array of molecules with anesthetic, immunosuppressive, vasodilatory, profibrinolytic, and anticoagulant properties (Mans and Neitz, 2004).
A transcript similar to longistatin, an EF-hand calcium binding protein, was overexpressed in ticks fed tick-susceptible cattle. The EF-hand calcium binding proteins modulate several crucial biochemical reactions within the cell and have a role in the blood clotting cascade, being secreted in some rare cases, as observed for the longistatin transcript identified by Anisuzzaman et al. (2012). The authors verified an overexpression of longistatin in salivary glands of the ixodide H. longicornis adults pre-engorged (96 h) and engorged (120 h) followed by an abrupt reduction in expression level with tick detachment, showing that longistatin clearly performs a vital function in the feeding process of the parasite through its anticoagulant action, resulting in fibrinogen hydrolysis and plasminogen activation. The post-transcriptional silencing of the longistatin gene in H. longicornis completely disrupted the ability of the parasite to form a blood pool and perform blood meals (Anisuzzaman et al., 2011).
In addition to the anticoagulant effect, longistatin has also been found to be able to modulate the inflammatory response of the host in a host-parasite interaction. The receptor for advanced glycation end products (RAGE) is highly expressed constitutively in different cell types of the skin and is responsible for immune activation mediation at inflammation sites. Anisuzzaman and Tsuji (2015) demonstrated that longistatin secreted in H. longicornis saliva binds to RAGE, modulating the host immune response and suppressing the inflammation associated with the lesion caused by the tick bite, thereby ensuring the successful acquisition of host blood. Immunization of mice with recombinant longistatin induced high levels of IgG protective antibodies against ticks, as well as reduced repletion of ticks by ~54%, post-engorgement body weight by ~11%, and nymph molting by ~34% (Anisuzzaman et al., 2012). H. longicornis treated with dsRNA failed to form a blood pool necessary for the proper blood meal, which led to a reduced engorged tick weight (Anisuzzaman et al., 2011).
Other binding Ca++-containing EF-hand motif molecules, similar to longistatin, have recently been identified from Strongyloides venezuelensis, the venestatin (Tsubokawa et al., 2017) and Aedes aegypt—the salivary factor LTRIN (Jin et al., 2018), which were upregulated promptly after larval invasion through the host's skin and during mosquito blood feeding, respectively.
A transcript similar to a serine carboxypeptidase (SCP) was upregulated in ticks fed on susceptible cattle compared to those fed on resistant cattle. SCPs are proteolytic enzymes that target serine in their catalytic activity and catalyze the hydrolysis of C-terminal residues in peptides and proteins (Anderson et al., 2008). In mammals, SCPs are involved in signaling events, such as the generation of bioactive peptides in the blood coagulation cascade (Turk, 2006), but little is known about SCPs in tick species. In Ixodes ricinus, enzymes as cathepsins B, C, and D, legumain, serine carboxypeptidase and leucine aminopeptidase were characterized as the main enzymes acting in the tick hemoglobinolytic system at midgut (Horn et al., 2009). Motobu et al. (2007) identified an H. longicornis SCP predominantly expressed in tick midgut upregulated in response to blood feeding, which has been described to play a role in hemoglobin degradation. SCPs were highly expressed in salivary glands and viscera from fully engorged Ixodes holocyclus female ticks (Rodriguez-Valle et al., 2018). The secretory/excretory proteome of helminths parasites identified SCPs that are part of a protein complex that regulates parasite-host interaction (Morassutti et al., 2012). Radulović et al. (2014) identified SCPs transcribed in the salivary gland of Amblyoma americanum that could act to regulate the interaction with the host.
Insect cells usually express a number of β-N-acetylhexosaminidases, which are important in various developmental stages during their life cycle, primarily in the turnover of the chitin exoskeleton (Hogenkamp et al., 2008). According to You and Fujisaki (2009), molecules associated with molt act to protect organisms against pathogen invasion, on peritrophic membrane control and other functions necessary during blood feeding and molting in ticks and may be candidates for parasite control. In our results, a DEG similar to a β-N-acetylhexosaminidase and upregulated in ticks fed on tick-susceptible cattle was found in salivary glands. Del Pino et al. (1998) inoculated anti-β-N-acetylhexosaminidase antibodies obtained against the enzyme purified from larvae of R. (B.) microplus into fully engorged, adult females and verified a decrease in the reproductive efficiency of R. (B.) microplus. Chitin is a polysaccharide that consists predominantly of unbranched chains of N-acetylglucosamine and is present together with a protein matrix in the cuticle of arthropods, including R. (B.) microplus (Hackman, 1975). Ticks periodically shed their old cuticles and resynthesize new ones, a process that is mediated by the elaboration of chitinase in the molting fluid that accumulates between the old cuticle and the epidermis (You and Fujisaki, 2009). Female ticks of the Ixodidae family, in general, increase their mass up to 100-fold during the 7–10 days feeding period and synthesize sufficient endocuticle to thicken their cuticle by the end of the slow phase of engorgement (Flynn and Kaufman, 2015); thus, the inhibition of enzymes involved in chitin metabolism during morphogenesis might cause serious tick damage (Del Pino et al., 1998).
A transcript similar to leucine aminopeptidase (LAP) was upregulated in ticks fed tick-susceptible cattle. Leucine aminopeptidases belong to a diverse group of the M17 family of Zn-metalloproteases and preferentially cleave a leucine residue at the N-terminus of the proteins and peptides (Maggioli et al., 2018). Metalloproteases, which have been found in tick saliva, salivary gland, ovary and midgut, play an important role in inflammation, immunomodulation, fibrinolysis, blood protein digestion, nociception, vitellogenesis, remodeling of extracellular matrix, and pathogen transmission (Ali et al., 2015). Leucine aminopeptidases have been identified in the midgut and salivary glands of engorged R. (B.) microplus (Kerlin and Hughes, 1992) and in the cytosol of cells of the midgut, salivary glands type II acini and epidermal cells from H. longicornis (Hatta et al., 2006). Overexpression of the midgut enzyme has been observed during blood feeding in H. longicornis, where the disruption of the gene encoding LAP through RNAi significantly delayed onset of egg-laying and reduced egg oviposition, suggesting that HlLAP plays a role as a blood digestive enzyme and affects tick fecundity (Hatta et al., 2007). In sheep, immunization with a leucine aminopeptidase purified from Fasciola hepatica resulted in high levels of animal protection against the endoparasite (Piacenza et al., 1999).
Ribosomal proteins play essential roles in cell growth and proliferation through the ribosome biogenesis process to translate mRNAs into proteins (Trainor and Merrill, 2014), and the assembly of a functional ribosome is vital for successful protein synthesis. In our study, two transcripts with similarity to ribosomal proteins were found among the DEGs in the salivary gland of R. (B.) microplus. The first transcript, a 60S acidic ribosomal protein P2, was upregulated in the salivary glands of ticks feeding on tick-susceptible cattle.
Phosphoproteins P1 and P2 form a complex with P0 protein to raise the eukaryotic ribosomal stalk. A number of ribosome-associated proteins were identified in female tissues from R. (B.) microplus, including components of the 40S and ribosomal subunits, as well as the 29S and mitochondrial ribosomal subunits (Stutzer et al., 2013), and in the salivary gland immune-proteome of A. americanum (Radulović et al., 2014). In the salivary gland transcriptome of Rhipicephalus appendiculatus, most represented pathways were ribosome, RNA transport, protein processing in endoplasmic reticulum, and spliceosome (de Castro et al., 2016). Ribosomal P proteins seem to play an important role in diseases related to infections caused by intracellular protozoan parasites. Rodríguez-Mallon et al. (2015) identified an immunogenic region of ribosomal protein P0 from Rhipicephalus spp. ticks and a synthetic 20 amino acid peptide from this sequence showed an efficacy of 96% as a vaccine against Rhipicephalus sanguineus in an immunization experiment with rabbits.
The second transcript, with similarity to 60S ribosomal protein L19 (RPL19), was upregulated in ticks feeding on tick-resistant cattle. Despite the essential role of the ribosome in protein synthesis, RPL19 is predicted to have an extra-protein translational function, and its overexpression in cell culture activated the cellular signaling pathway of the UPR (unfolded protein response) (Hong et al., 2014). The UPR pathway is conserved from yeast to humans (Zhang et al., 2016) and activated in response to an overloading of unfolded or misfolded peptides, and its action can either have a protective effect, with the change of protein synthesis rate decreasing peptide load into the endoplasmic reticulum or inducing cell death with apoptotic characteristics (Kim et al., 2008; Hong et al., 2014). The RPL19 gene was upregulated in ticks fed tick-resistant cattle, which indicates that cell death was the main pathway activated to salivary gland cells during host-parasite interactions. In Jurkat cells, RPL19 protein expression was decreased by heat stress, although there was no significant change in mRNA expression (Zhang et al., 2016).
Phosphorylase kinase (PhK) is a holoenzyme that activates glycogen phosphorylase (Brushia and Walsh, 1999) which, in turn, catalyzes the hydrolysis of glycogen to generate glucose-1-phosphate. In ticks fed on tick-susceptible cattle, an upregulation in a transcript similar to PhK was observed.
Genes involved in stress response include molecular chaperones, such as members of the heat shock protein gene family, as already mentioned, in addition to antioxidative enzymes (e.g., catalase, superoxide dismutases, and glutathione-S-transferases) and enzymes of carbohydrate metabolism (e.g., glycogen phosphorylase and phosphofructokinase) (Barat et al., 2016). In mammals, glycogen metabolism is profoundly affected by stress, with glycogen content in peripheral tissues decreasing during stress because of increased glycogenolysis and decreased glycogen synthesis (Van Cromphaut, 2009). To date, few studies have investigated tick glycogen metabolism. However, in a study on arthropod thermal tolerance, a reduction in glycogen content was observed in D. melanogaster with a sudden increase in temperature from 25 to 41.2–41.3°C with a ramping rate of 0.1°C min−1 (Overgaard et al., 2012). As appears to have occurred with HSPI and NELF transcripts, the upregulation of PHKA in ticks feeding on tick-susceptible cattle may be related to the stress condition imposed by blood feeding.
A transcript that encodes a conserved hypothetical protein (XP_002414744.1) with cullin-binding and a UBA_DCNL2 domain was upregulated in ticks feeding on tick-susceptible cattle. This protein functions as an E3 ligase, promoting cullin neddylation by binding to cullins through conserved interaction surfaces on each protein (Kurz et al., 2008). Neddylation is a post-translational modification process analogous to ubiquitination, where neddylated proteins are modified through the closest relative to ubiquitin—NEDD8 protein, which attaches to a lysine residue of the cullin scaffold proteins (Sakata et al., 2007; Rabut and Peter, 2008). Neddylation is important for a number of biological processes and is required for the regulation of a multifunctional transcription factor, NF-κB, which is crucial in immune-response and apoptotic pathways (Cajee et al., 2012). The gene upregulation verified in ticks fed on tick-susceptible cattle in our study corroborates the role of pathways related to cellular proteolysis, as already mentioned in the case of upregulation of HSP1, GGT, and downregulation of NELF, in the maintenance of the susceptible host-tick interaction.
In vertebrates, mucins are abundant in the lungs and digestive tract, where they provide lubrication and protection of the epithelium against physical damage and pathogens (Hollingsworth and Swanson, 2004). Mucins are either secreted and gel-forming or attached to the membrane by special cleavable transmembrane domains (Syed et al., 2008). A mucin-like family has been identified in insects, and its family members are distributed in salivary glands, midgut, and Malpighian tubules (Barry et al., 1999; Syed et al., 2008; Hegedus et al., 2009). In our study, a transcript similar to a mucin-17-like isoform X3 was upregulated in ticks fed on crossbreed cattle. In the hemipteran Nilaparvata lugens, the rice brown planthopper (BPH), a mucin-like transcript, was upregulated when the insects were transferred from a susceptible rice variety to a resistant one (Huang et al., 2017). The authors suggested that the elevated mucin production might enable BPH to cope with the stress of the defense responses or help BPH to suppress the defenses of the resistant plant (Huang et al., 2017). An upregulation in a mucin-17-like isoform X3 transcript was also observed in Diuraphis noxia fed on an aphid-resistant wheat plant in comparison to non-resistant plants (Sinha et al., 2016). In mice immunized with Anopheles gambiae midgut-bound mucin cDNA, increased mortality was observed among mosquitoes fed immunized mice compared to those fed control mice (Foy et al., 2003).
Mucin and mucin-like proteins have been found in R. (B.) microplus (Maruyama et al., 2017) and the sialotranscriptomes of other tick species (Karim et al., 2011; Díaz-Martín et al., 2013; Radulović et al., 2014; Tan et al., 2015; Ong et al., 2016; Antunes et al., 2018), and they may function in tick feeding by coating the chitinous feeding mouthparts or the feeding lesion (Francischetti, 2010). In 1998, an antigen purified from R. (B.) microplus and named BMA7, exhibiting similarity to vertebrate mucins, induced partial immunity against tick infestation in cattle (Mckenna et al., 1998).
Innexins are transmembrane proteins that form gap junction channels and hemichannels in invertebrates, including arthropods (Richards et al., 2015; Güiza et al., 2018). Gap junctions allow endocrine signals to be rapidly shared among adjacent cells within a tissue by mediating the direct transport of ions, small molecules, and second messengers between them (Phelan and Starich, 2001). Pharmacological inhibitors of gap junctions, such as carbenoxolone, mefloquine, and meclofenamic acids, have been cited as potential insecticides in arthropod vectors, such as Aedes aegypti (Calkins and Piermarini, 2015). In our study, a transcript similar to innexin was downregulated in the salivary glands of ticks fed on resistant cattle. A transcript with similarity to innexin was also expressed in the salivary glands of the female tick A. americanum during blood feeding (Aljamali et al., 2009) and in the proteome of the saliva of the tick O. moubata (Díaz-Martín et al., 2013).
The gene that encodes the predicted secreted protein Na+/dicarboxylate, Na+/tricarboxylate, and phosphate transporter, was upregulated in salivary glands of ticks fed on susceptible cattle. Analyzing transcriptome changes in Eriocheir sinensis larvae after desalination, Hui et al. (2014) identified transcripts of many genes involved in ion transport processes, including the Na+/dicarboxylate, Na+/tricarboxylate, and phosphate transporter, to which it was attributed a potential role in osmoregulation.
A protein predicted as secreted, the phosphoinositol 4-phosphate adaptor protein, was upregulated in ticks feeding on resistant cattle. Four phosphate-adaptor proteins 1 and 2 (FAPP1 and FAPP2) are proteins that localize to the trans-Golgi network (TGN) on nascent carriers and interact with phosphatidylinositol-4-phosphate and the small GTPase ADP-ribosylation factor (ARF) through their plekstrin homology domain (Godi et al., 2004). Phosphoinositol 4-phosphate adaptor proteins are involved in lipid transport (Oleaga et al., 2017) and control the formation and fission of post-Golgi vesicles and regulate secretory transport from the TGN to the plasma membrane, and displacement or knockdown of FAPPs inhibits cargo transfer to the plasma membrane (Godi et al., 2004). In Ornithodoros moubata ticks, phosphoinositol 4-phosphate adaptor protein transcripts were upregulated in the midgut after blood feeding in comparison with non-fed ticks (Oleaga et al., 2017). The phosphatidylinositol-4-phosphate, in turn, is a minor plasma membrane phospholipid component of a signal transduction pathway in tick salivary glands (McSwain et al., 1989), a key regulator of membrane transport required for the formation of transport carriers from the TGN. ARF is a Ras-related GTP-binding protein that regulates the reversible binding of cytosolic coat proteins to Golgi membranes (Donaldson and Klausner, 1994).
A putative monocarboxylate transporter was identified on the A. americanum tick saliva immunoproteome from fed adult females (Radulović et al., 2014) and overexpressed in R. (B.) microplus engorged females treated with ubiquitin-63E dsRNA (Lew-Tabor et al., 2011). Monocarboxylate transporters catalyze rapid transport across the plasma membrane of many monocarboxylates, such as lactate and pyruvate (Lew-Tabor et al., 2011). A transcript similar to a monocarboxylate transporter, predicted as a secreted protein, was upregulated in ticks fed on tick-susceptible cattle.
A secreted cysteine-rich protein containing a trypsin inhibitor-like (TIL) domain was upregulated in the salivary glands of ticks feeding on tick-susceptible cattle. This family comprises chymotrypsin, elastase and trypsin inhibitors (Sasaki et al., 2008), although many extracellular proteins also contain multiple TIL domains (Wang et al., 2015). TIL domain-containing protein family members have been found ubiquitously in blood-feeding insect and tick sialomes (Karim and Adamson, 2012; Maruyama et al., 2017), and in fungal-pathogen interactions, members of this family are frequently associated with host adaptation or specialization (Schulze-Lefert and Panstruga, 2011).
In R. (B.) microplus, a member of this family, known as ixodidin, was purified from the hemocytes and characterized as an antimicrobial peptide, which affected the growth of Micrococcus luteus and Escherichia coli and presented inhibitory activity (Fogaça et al., 2006). BmSI-7, a protein from R. (B.) microplus belonging to the trypsin inhibitor-like cysteine-rich domain family showed strong inhibitory activity toward elastase, which participates in the inflammatory response and is involved in injury caused by tick fixation on bovines (Sasaki et al., 2008). A secreted cysteine-rich protein containing the TIL domain was upregulated in the proteome of fed Amblyomma sculptum, induced by blood feeding, compared with non-fed ticks (Esteves et al., 2017). In R. (B.) microplus, TIL domain-containing proteins were also upregulated in the fully engorged female proteome in comparison with partially engorged females (Tirloni et al., 2014).
A transcript similar to the putative defense protein 3 containing a reeler domain, predicted to be a secreted protein, was upregulated in ticks feeding on tick-susceptible cattle. A putative defense protein 3 was also upregulated in the proteome of a whitefly infected by two begomoviruses, the tomato yellow leaf curl virus (TYLCV) and the papaya leaf curl China virus (PaLCuCNV), as a defense response against the viruses (Zhao et al., 2019). Defense proteins with a reeler domain have been shown to have an important role in innate immune responses in a variety of insects (Bao et al., 2011; Arp et al., 2016). In I. scapularis, a predicted secreted protein with a reeler domain—PIXR—was induced upon feeding and upregulated in B. burgdorferi-infected tick guts, favoring colonization by the bacteria (Narasimhan et al., 2017).
In our results, a transcript similar to an F-actin-uncapping protein, LRRC16A isoform X2 (LRCC16A) with a CARMIL_C domain, a putative secreted protein, was upregulated in salivary glands from ticks feeding on tick-resistant cattle. Actins are the major constituents of the actin cytoskeleton and are essential for cell adhesion, migration, mechanosensing, and contractility in muscle and non-muscle tissues (Simiczyjew et al., 2017; Vedula and Kashina, 2018). Actin filaments grow and shrink by addition and loss, respectively, of actin monomers at the ends of filaments (Stark et al., 2017), and such regulation affects a wide range of cell processes, including development, differentiation, immunity, and inflammation (Marcos-Ramiro et al., 2014). Actin polymerization occurs primarily through elongation at the filament barbed end, and the elongation continues until the barbed end is capped by a capping protein (CP) (Yang et al., 2005). Regulation of barbed-end capping occurs by binding of the inhibitor factors to the filament, thereby protecting it from CP, by binding to CP and inhibiting its capping activity or by uncapping (Yang et al., 2005). CARMILs are multidomain proteins that regulate the actin-binding activity of CP binding directly to it and induce a conformational change that decreases its actin-capping activity (Stark et al., 2017).
Our results help to characterize cattle tick salivary gland gene expression in both susceptible and resistant hosts and suggest new putative targets for infestation control, as those genes involved in stress response mechanism during blood feeding. A possible coordinated regulation targeting a small heat shock protein and a negative elongation factor B-like genes, the latter expressed in order to maintain heat shock gene expression increased during blood feeding; along with upregulation of a 26S proteasome subunit and calnexin, other chaperone, shed light to the role of this mechanism in maintaining tick feeding. Other interactions, as the one described, are under analysis, and may evince important new targets to vaccines development.
The datasets generated for this study can be found in the NCBI SRA database under accessions SAMN13636118 and SAMN13636119 from BioProject PRJNA596777 (http://www.ncbi.nlm.nih.gov/bioproject/596777).
The animal study was reviewed and approved by Ethics Statement. All experimental procedures were approved by Embrapa Beef Cattle's Ethics Committee on Animal Use according to Protocol 008/2014; coordinated by Vanessa Felipe Souza. e-mail: dmFuZXNzYS5mZWxpcGVAZW1icmFwYS5icg==.
RA and RC conceived and designed the study. MG and RC were responsible for the tick rearing, cattle artificial infestation, tick collection, and RNA isolation. PG and AN performed the data analysis. PG, AN, and RA wrote the manuscript. JF critically revised the manuscript. All authors edited and approved the final manuscript.
This work was supported by funds from the Brazilian Agricultural Research Corporation-Embrapa, grants 01.11.07.002.03.00.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank the Foundation for Support to the Development of Education, Science and Technology of the State of Mato Grosso do Sul (FUNDECT/MS), the Government of the state of Mato Grosso do Sul, the Brazilian Federal Agency for Support and Evaluation of Graduate Education (Capes), the National Council for Scientific and Technological Development (CNPq), and Embrapa Bioinformatic Multi-user Laboratory (LMB) for providing support and computing resources.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2019.00477/full#supplementary-material
Accaoui, M. J., Enoiu, M., Mergny, M., Masson, C., Dominici, S., Wellman, M., et al. (2000). Gamma-glutamyltranspeptidase-dependent glutathione catabolism results in activation of NF-kB. Biochem. Biophys. Res. Commun. 276, 1062–1067. doi: 10.1006/bbrc.2000.3585
Ali, A., Khan, S., Ali, I., Karim, S., da Silva Vaz, I. Jr., and Termignoni, C. (2015). Probing the functional role of tick metalloproteases. Physiol. Entomol. 40, 177–188. 10.1111/phen.12104 doi: 10.1111/phen.12104
Aljamali, M. N., Hern, L., Kupfer, D., Downard, S., So, S., Roe, B. A., et al. (2009). Transcriptome analysis of the salivary glands of the female tick Amblyomma americanum (Acari: Ixodidae). Insect Mol. Biol. 18, 129–154. doi: 10.1111/j.1365-2583.2009.00863.x
Anderson, J. M., Sonenshine, D. E., and Valenzuela, J. G. (2008). Exploring the mialome of ticks: an annotated catalogue of midgut transcripts from the hard tick, Dermacentor variabilis (Acari: Ixodidae). BMC Genomics 9:552. doi: 10.1186/1471-2164-9-552
Andrade, H. M., Murta, S. M., Chapeaurouge, A., Perales, J., Nirdé, P., and Romanha, A. J. (2008). Proteomic analysis of Trypanosoma cruzi resistance to benznidazole. J. Proteome Res. 7, 2357–2367. doi: 10.1021/pr700659m
Anisuzzaman, M., and Tsuji, N. (2015). Longistatin in tick-saliva targets RAGE. Oncotarget 6, 35133–35134. doi: 10.18632/oncotarget.6032
Anisuzzaman, M. K. I., Alim, M. A., Miyoshi, T., Hatta, T., Yamaji, K., Matsumoto, Y., et al. (2011). Longistatin, a plasminogen activator, is key to the availability of blood-meals for ixodid ticks. PLoS Pathog. 7:e1001312. doi: 10.1371/journal.ppat.1001312
Anisuzzaman, M. K. I., Alim, M. A., Miyoshi, T., Hatta, T., Yamaji, K., Matsumoto, Y., et al. (2012). Longistatin is an unconventional serine protease and induces protective immunity against tick infestation. Mol. Biochem. Parasitol. 182, 45–53. doi: 10.1016/j.molbiopara.2011.12.002
Antunes, S., Couto, J., Ferrolho, J., Rodrigues, F., Nobre, J., Santos, A. S., et al. (2018). Rhipicephalus bursa sialotranscriptomic response to blood feeding and Babesia ovis infection: identification of candidate protective antigens. Front. Cell. Infect. Microbiol. 8:116. doi: 10.3389/fcimb.2018.00116
Arp, A. P., Hunter, W. B., and Pelz-Stelinski, K. S. (2016). Annotation of the Asian citrus psyllid genome reveals a reduced innate immune system. Front. Physiol. 7:570. doi: 10.3389/fphys.2016.00570
Bao, Y. Y., Xue, J., Wu, W. J., Wang, Y., Lv, Z. Y., and Zhang, C. X. (2011). An immune-induced reeler protein is involved in the Bombyx mori melanization cascade. Insect Biochem. Mol. Biol. 41, 696–706. doi: 10.1016/j.ibmb.2011.05.001
Barat, A., Sahoo, P. K., Kumar, R., Goel, C., and Singh, A. K. (2016). Transcriptional response to heat shock in liver of snow trout (Schizothorax richardsonii)–a vulnerable Himalayan Cyprinid fish. Funct. Integr. Genomics 16, 203–213. doi: 10.1007/s10142-016-0477-0
Barry, M. K., Triplett, A. A., and Christensen, A. C. (1999). A peritrophin-like protein expressed in the embryonic tracheae of Drosophila melanogaster. Insect Biochem. Mol. Biol. 29, 319–327. doi: 10.1016/S0965-1748(99)00004-1
Benoit, J. B., Lazzari, C. R., Denlinger, D. L., and Lahondère, C. (2019). Thermoprotective adaptations are critical for arthropods feeding on warm-blooded hosts. Curr. Opin. Insect Sci. 34, 7–11. doi: 10.1016/j.cois.2019.02.003
Benoit, J. B., Lopez-Martinez, G., Patrick, K. R., Phillips, Z. P., Krause, T. B., and Denlinger, D. L. (2011). Drinking a hot blood meal elicits a protective heat shock response in mosquitoes. PNAS 108, 8026–8029. doi: 10.1073/pnas.1105195108
Bergman, D. K., Palmer, M. J., Caimano, M. J., Radolf, J. D., and Wikel, S. K. (2000). Isolation and molecular cloning of a secreted immunosuppressant protein from Dermacentor andersoni salivary gland. J. Parasitol. 86, 516–525. doi: 10.2307/3284866
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Brushia, R. J., and Walsh, D. A. (1999). Phosphorylase kinase: the complexity of its regulation is reflected in the complexity of its structure. Front. Biosci. 4, D618–D641. doi: 10.2741/Brushia
Buchfink, B., Xie, C., and Huson, D. H. (2015). Fast and sensitive protein alignment using DIAMOND. Nat. Methods 12, 59–60. doi: 10.1038/nmeth.3176
Cajee, U. F., Hull, R., and Ntwasa, M. (2012). Modification by ubiquitin-like proteins: significance in apoptosis and autophagy pathways. Int. J. Mol. Sci. 13, 11804–11831. doi: 10.3390/ijms130911804
Calkins, T. L., and Piermarini, P. M. (2015). Pharmacological and genetic evidence for gap junctions as potential new insecticide targets in the yellow fever mosquito, Aedes aegypti. PLoS ONE 10:e0137084. doi: 10.1371/journal.pone.0137084
Choudhury, R. R., Hyvola, N., and Lowe, M. (2005). Phosphoinositides and membrane traffic at the trans-Golgi network. Biochem. Soc. Symp. 72, 31–38. doi: 10.1042/bss0720031
Coux, O., Tanaka, K., and Goldberg, A. L. (1996). Structure and functions of the 20S and 26S proteasomes. Annu. Rev. Biochem. 65, 801–847. doi: 10.1146/annurev.bi.65.070196.004101
Csordas, B. G., Cunha, R. C., Garcia, M. V., da Silva, S. S., Leite, F. L., and Andreotti, R. (2018). Molecular characterization of the recombinant protein RmLTIBmCG-LTB: protective immunity against Rhipicephalus (Boophilus) microplus. PLoS ONE 13:e0191596. doi: 10.1371/journal.pone.0191596
de Castro, M. H., De Klerk, D., Pienaar, R., Latif, A. A., Rees, D. J. G., and Mans, B. J. (2016). De novo assembly and annotation of the salivary gland transcriptome of Rhipicephalus appendiculatus male and female ticks during blood feeding. Ticks Tick Borne Dis. 7, 536–548. doi: 10.1016/j.ttbdis.2016.01.014
de Lima, M. R., Ferreira, C. S., de Freitas, D. J., Valenzuela, J. G., and Masuda, A. (2002). Cloning and partial characterization of a Boophilus microplus (Acari: Ixodidae) glutathione S-transferase. Insect Biochem. Mol. Biol. 32, 747–754. doi: 10.1016/S0965-1748(01)00157-6
Del Pino, F. A., Brandelli, A., Gonzales, J. C., Henriques, J. A., and Dewes, H. (1998). Effect of antibodies against beta-N-acetylhexosaminidase on reproductive efficiency of the bovine tick Boophilus microplus. Vet. Parasitol. 79, 247–255. doi: 10.1016/S0304-4017(98)00154-X
Díaz-Martín, V., Manzano-Román, R., Valero, L., Oleaga, A., Encinas-Grandes, A., and Pérez-Sánchez, R. (2013). An insight into the proteome of the saliva of the argasid tick Ornithodoros moubata reveals important differences in saliva protein composition between the sexes. J. Proteomics 80, 216–235. doi: 10.1016/j.jprot.2013.01.015
Donaldson, J. G., and Klausner, R. D. (1994). ARF: a key regulatory switch in membrane traffic and organelle structure. Curr. Opin. Cell. Biol. 6, 527–532. doi: 10.1016/0955-0674(94)90072-8
Esteves, E., Maruyama, S. R., Kawahara, R., Fujita, A., Martins, L. A., Righi, A. A., et al. (2017). Analysis of the salivary gland transcriptome of unfed and partially fed Amblyomma sculptum ticks and descriptive proteome of the saliva. Front. Cell. Infect. Microbiol. 7:476. doi: 10.3389/fcimb.2017.00476
Flynn, P. C., and Kaufman, W. R. (2015). Mechanical properties of the cuticle of the tick Amblyomma hebraeum (Acari: Ixodidae). J. Exp. Biol. 218, 2806–2814. doi: 10.1242/jeb.123919
Fogaça, A. C., Almeida, I. C., Eberlin, M. N., Tanaka, A. S., Bulet, P., and Daffre, S. (2006). Ixodidin, a novel antimicrobial peptide from the hemocytes of the cattle tick Boophilus microplus with inhibitory activity against serine proteinases. Peptides 27, 667–674. doi: 10.1016/j.peptides.2005.07.013
Foy, B. D., Magalhaes, T., Injera, W. E., Sutherland, I., Devenport, M., Thanawastien, A., et al. (2003). Induction of mosquitocidal activity in mice immunized with Anopheles gambiae midgut cDNA. Infect. Immun. 71, 2032–2040. doi: 10.1128/IAI.71.4.2032-2040.2003
Francischetti, I. M. B. (2010). Platelet aggregation inhibitors from hematophagous animals. Toxicon 56, 1130–1144. doi: 10.1016/j.toxicon.2009.12.003
Franzin, A. M., Maruyama, S. R., Garcia, G. R., Oliveira, R. P., Ribeiro, J. M., Bishop, R., et al. (2017). Immune and biochemical responses in skin differ between bovine hosts genetically susceptible and resistant to the cattle tick Rhipicephalus microplus. Parasit. Vectors 10:51. doi: 10.1186/s13071-016-1945-z
Gallastegui, N., and Groll, M. (2010). The 26S proteasome: assembly and function of a destructive machine. Trends Biochem. Sci. 35, 634–642. doi: 10.1016/j.tibs.2010.05.005
Ghosh, S., Khan, M. H., and Ahmed, N. (1999). Cross-bred cattle protected against Hyalomma anatolicum anatolicum by larval antigens purified by immunoaffinity chromatography. Trop. Anim. Health Prod. 31, 263–273. doi: 10.1023/A:1005218821889
Ghosh, S. K. B., Missra, A., and Gilmour, D. S. (2011). Negative elongation factor accelerates the rate at which heat shock genes are shut off by facilitating dissociation of heat shock factor. Mol. Cell. Biol. 31, 4232–4243. doi: 10.1128/MCB.05930-11
Godi, A., Di Campli, A., Konstantakopoulos, A., Di Tullio, G., Alessi, D. R., Kular, G. S., et al. (2004). FAPPs control Golgi-to-cell-surface membrane traffic by binding to ARF and PtdIns (4) P. Nat. Cell Biol. 6, 393–404. doi: 10.1038/ncb1119
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., et al. (2011). Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 29, 644–652. doi: 10.1038/nbt.1883
Grisi, L., Leite, R. C., Martins, J. R. S., Barros, A. T. M., Andreotti, R., Cançado, P. H. D., et al. (2014). Reassessment of the potential economic impact of cattle parasites in Brazil. Rev. Bras. Parasitol. Vet. 23, 150–156. doi: 10.1590/S1984-29612014042
Guérin, R., Arseneault, G., Dumont, S., and Rokeach, L. A. (2008). Calnexin is involved in apoptosis induced by endoplasmic reticulum stress in the fission yeast. Mol. Biol. Cell 19, 4404–4420. doi: 10.1091/mbc.e08-02-0188
Guilfoile, P. G., and Packila, M. (2004). Identification of four genes expressed by feeding, female Ixodes scapularis including three with sequence similarity to previously recognized genes. Exp. Appl. Acarol. 32, 103–110. doi: 10.1023/B:APPA.0000018197.63153.58
Güiza, J., Barria, I., Saez, J. C., and Vega, J. L. (2018). Innexins: expression, regulation and functions. Front. Physiol. 9:1414. doi: 10.3389/fphys.2018.01414
Haas, B. J., Papanicolaou, A., Yassour, M., Grabherr, M., Blood, P. D., Bowden, J., et al. (2013). De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 8, 1494–1512. doi: 10.1038/nprot.2013.084
Hackman, R. H. (1975). Expanding abdominal cuticle in the bug Rhodnius and the tick Boophilus. J. Insect Physiol. 21, 1613–1623. doi: 10.1016/0022-1910(75)90199-7
Hatta, T., Kazama, K., Miyoshi, T., Umemiya, R., Liao, M., Inoue, N., et al. (2006). Identification and characterisation of a leucine aminopeptidase from the hard tick Haemaphysalis longicornis. Int. J. Parasitol. 36, 1123–1132. doi: 10.1016/j.ijpara.2006.05.010
Hatta, T., Umemiya, R., Liao, M., Gong, H., Harnnoi, T., Tanaka, M., et al. (2007). RNA interference of cytosolic leucine aminopeptidase reduces fecundity in the hard tick, Haemaphysalis longicornis. Parasitol. Res. 100, 847–854. doi: 10.1007/s00436-006-0336-3
Hegedus, D., Erlandson, M., Gillott, C., and Toprak, U. (2009). New insights into peritrophic matrix synthesis, architecture, and function. Annu. Rev. Entomol. 54, 285–302. doi: 10.1146/annurev.ento.54.110807.090559
Hernandez, E. P., Kusakisako, K., Talactac, M. R., Galay, R. L., Hatta, T., Fujisaki, K., et al. (2018). Glutathione S-transferases play a role in the detoxification of flumethrin and chlorpyrifos in Haemaphysalis longicornis. Parasit. Vectors 11:460. doi: 10.1186/s13071-018-3044-9
Hewetson, R. W. (1972). The inheritance of resistance by cattle to cattle tick. Aust. Vet. J. 48, 299–303. doi: 10.1111/j.1751-0813.1972.tb05161.x
Higa, L. O. S., Garcia, M. V., Rodrigues, V. S., Bonatte, J. P., Piña, F. T. B., Barros, J. C., et al. (2019). Effects of cypermethrin, chlorpyrifos and piperonyl butoxide-based pour-on and spray acaricides on controlling the tick Rhipicephalus microplus. Syst. Appl. Acarol. 24, 278–286. doi: 10.11158/saa.24.2.10
Hogenkamp, D. G., Arakane, Y., Kramer, K. J., Muthukrishnan, S., and Beeman, R. W. (2008). Characterization and expression of the β-N-acetylhexosaminidase gene family of Tribolium castaneum. Insect Biochem. Mol. Biol. 38, 478–489. doi: 10.1016/j.ibmb.2007.08.002
Hollingsworth, M. A., and Swanson, B. J. (2004). Mucins in cancer: protection and control of the cell surface. Nat. Rev. Cancer 4, 45–60. doi: 10.1038/nrc1251
Hong, M., Kim, H., and Kim, I. (2014). Ribosomal protein L19 overexpression activates the unfolded protein response and sensitizes MCF7 breast cancer cells to endoplasmic reticulum stress-induced cell death. Biochem. Biophys. Res. Commun. 450, 673–678. doi: 10.1016/j.bbrc.2014.06.036
Horn, M., Nussbaumerová, M., Šanda, M., Kovárová, Z., Srba, J., Franta, Z., et al. (2009). Hemoglobin digestion in blood-feeding ticks: mapping a multipeptidase pathway by functional proteomics. Chem. Biol. 16, 1053–1063. doi: 10.1016/j.chembiol.2009.09.009
Hovius, J. W. R., Levi, M., and Fikrig, E. (2008). Salivating for knowledge: potential pharmacological agents in tick saliva. PLoS Med. 5:e43. doi: 10.1371/journal.pmed.0050043
Huang, H. J., Liu, C. W., Xu, H. J., Bao, Y. Y., and Zhang, C. X. (2017). Mucin-like protein, a saliva component involved in brown planthopper virulence and host adaptation. J. Insect Physiol. 98, 223–230. doi: 10.1016/j.jinsphys.2017.01.012
Hui, M., Liu, Y., Song, C., Li, Y., Shi, G., and Cui, Z. (2014). Transcriptome changes in Eriocheir sinensis megalopae after desalination provide insights into osmoregulation and stress adaption in larvae. PLoS ONE 9:e114187. doi: 10.1371/journal.pone.0114187
Hussein, N. A., Shahein, Y. E., El-Hakim, A. E., Abouelella, A. M., Guneidy, R. A., and Hamed, R. R. (2014). Molecular cloning of Ra-sHSPI, a novel member of the HSP20 family from Rhipicephalus annulatus salivary glands. Int. J. Biol. Macromol. 67, 7–15. doi: 10.1016/j.ijbiomac.2014.02.057
Jin, L., Guo, X., Shen, C., Hao, X., Sun, P., Li, P., et al. (2018). Salivary factor LTRIN from Aedes aegypti facilitates the transmission of Zika virus by interfering with the lymphotoxin-β receptor. Nat. Immunol. 19, 342–353. doi: 10.1038/s41590-018-0063-9
Karim, S., and Adamson, S. W. (2012). “RNA interference in ticks: a functional genomics tool for the study of physiology,” in Advances in Insect Physiology, ed E. L. Jockusch (London: Elsevier), 119–154. doi: 10.1016/B978-0-12-387680-5.00004-5
Karim, S., Singh, P., and Ribeiro, J. M. (2011). A deep insight into the sialotranscriptome of the gulf coast tick, Amblyomma maculatum. PLoS ONE 6:e28525. doi: 10.1371/journal.pone.0028525
Kerlin, R. L., and Hughes, S. (1992). Enzymes in saliva from four parasitic arthropods. Med. Vet. Entomol. 6, 121–126. doi: 10.1111/j.1365-2915.1992.tb00587.x
Kim, I., Xu, W., and Reed, J. C. (2008). Cell death and endoplasmic reticulum stress: disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 7, 1013–1030. doi: 10.1038/nrd2755
Kurz, T., Chou, Y. C., Willems, A. R., Meyer-Schaller, N., Hecht, M. L., Tyers, M., et al. (2008). Dcn1 functions as a scaffold-type E3 ligase for cullin neddylation. Mol. Cell 29, 23–35. doi: 10.1016/j.molcel.2007.12.012
Langmead, B., and Salzberg, S. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Lewis, L. A., Radulović, Ž. M., Kim, T. K., Porter, L. M., and Mulenga, A. (2015). Identification of 24 h Ixodes scapularis immunogenic tick saliva proteins. Ticks Tick Borne Dis. 6, 424–434. doi: 10.1016/j.ttbdis.2015.03.012
Lew-Tabor, A. E., Kurscheid, S., Barrero, R., Gondro, C., Moolhuijzen, P. M., Valle, M. R., et al. (2011). Gene expression evidence for off-target effects caused by RNA interference-mediated gene silencing of Ubiquitin-63E in the cattle tick Rhipicephalus microplus. Int. J. Parasitol. 41, 1001–1014. doi: 10.1016/j.ijpara.2011.05.003
Maggioli, G., Rinaldi, G., Giaudrone, I., Berasain, P., Tort, J. F., Brindley, P. J., et al. (2018). Expression, purification and characterization of two leucine aminopeptidases of the blood fluke, Schistosoma mansoni. Mol. Biochem. Parasitol. 219, 17–23. doi: 10.1016/j.molbiopara.2017.11.006
Mans, B. J., and Neitz, A. W. (2004). Adaptation of ticks to a blood-feeding environment: evolution from a functional perspective. Insect Biochem. Mol. Biol. 34, 1–17. doi: 10.1016/j.ibmb.2003.09.002
Marcos-Ramiro, B., García-Weber, D., and Millán, J. (2014). TNF-induced endothelial barrier disruption: beyond actin and Rho. Thromb. Haemost. 112, 1088–1102. doi: 10.1160/th14-04-0299
Maruyama, S. R., Garcia, G. R., Teixeira, F. R., Brandão, L. G., Anderson, J. M., Ribeiro, J. M., et al. (2017). Mining a differential sialotranscriptome of Rhipicephalus microplus guides antigen discovery to formulate a vaccine that reduces tick infestations. Parasit. Vectors 10:206. doi: 10.1186/s13071-017-2136-2
Mattioli, R. C., Bah, M., Faye, J., Kora, S., and Cassama, M. (1993). A comparison of field tick infestation on N'Dama, Zebu and N'Dama × Zebu crossbred cattle. Vet. Parasitol. 47, 139–148. doi: 10.1016/0304-4017(93)90184-O
Mckenna, R. V., Riding, G. A., Jarmey, J. M., Pearson, R. D., and Willadsen, P. (1998). Vaccination of cattle against the Boophilus microplus using a mucin-like membrane glycoprotein. Parasite Immunol. 20, 325–336. doi: 10.1046/j.1365-3024.1998.00149.x
McSwain, J. L., Tucker, J. S., Essenberg, R. C., and Sauer, J. R. (1989). Brain factor induced formation of inositol phosphates in tick salivary glands. Insect Biochem. 19, 343–349. doi: 10.1016/0020-1790(89)90038-3
Min, X. J. (2010). Evaluation of computational methods for secreted protein prediction in different eukaryotes. J. Proteomics Bioinform. 3, 143–147. doi: 10.4172/jpb.1000133
Morassutti, A. L., Levert, K., Pinto, P. M., da Silva, A. J., Wilkins, P., and Graeff-Teixeira, C. (2012). Characterization of Angiostrongylus cantonensis excretory–secretory proteins as potential diagnostic targets. Exp. Parasitol. 130, 26–31. doi: 10.1016/j.exppara.2011.10.003
Motobu, M., Tsuji, N., Miyoshi, T., Huang, X., Islam, M. K., Alim, M. A., et al. (2007). Molecular characterization of a blood-induced serine carboxypeptidase from the ixodid tick Haemaphysalis longicornis. FEBS J. 274, 3299–3312. doi: 10.1111/j.1742-4658.2007.05852.x
Mulenga, A., and Erikson, K. (2011). A snapshot of the Ixodes scapularis degradome. Gene 482, 78–93. doi: 10.1016/j.gene.2011.04.008
Nandi, A., Singh, H., and Singh, N. K. (2015). Esterase and glutathione S-transferase levels associated with synthetic pyrethroid resistance in Hyalomma anatolicum and Rhipicephalus microplus ticks from Punjab, India. Exp. Appl. Acarol. 66, 141–157. doi: 10.1007/s10493-015-9884-5
Narasimhan, S., Schuijt, T. J., Abraham, N. M., Rajeevan, N., Coumou, J., Graham, M., et al. (2017). Modulation of the tick gut milieu by a secreted tick protein favors Borrelia burgdorferi colonization. Nat. Commun. 8:184. doi: 10.1038/s41467-017-00208-0
Oleaga, A., Obolo-Mvoulouga, P., Manzano-Román, R., and Pérez-Sánchez, R. (2017). A proteomic insight into the midgut proteome of Ornithodoros moubata females reveals novel information on blood digestion in argasid ticks. Parasit. Vectors 10:366. doi: 10.1186/s13071-017-2300-8
Ong, C. T., Rodriguez-Valle, M., Barrero, R. A., Hunter, A., Bellgard, M. I., and Lew-Tabor, A. E. (2016). Exploring the transcriptomic data of the Australian paralysis tick, Ixodes holocyclus. GSTF J. Vet. Sci. 3. doi: 10.5176/2345-7880_3.1.14
Overgaard, J., Kristensen, T. N., and Sørensen, J. G. (2012). Validity of thermal ramping assays used to assess thermal tolerance in arthropods. PLoS ONE 7:e32758. doi: 10.1371/journal.pone.0032758
Pereira, M. H., Paim, R. M., Lahondère, C., and Lazzari, C. R. (2017). “Heat shock proteins and blood-feeding in arthropods,” in Heat Shock Proteins in Veterinary Medicine and Sciences, eds A. A. A. Asea and P. Kaur (Cham: Springer), 349–359. doi: 10.1007/978-3-319-73377-7_13
Phelan, P., and Starich, T. A. (2001). Innexins get into the gap. Bioessays 23, 388–396. doi: 10.1002/bies.1057
Piacenza, L., Acosta, D., Basmadjian, I., Dalton, J. P., and Carmona, C. (1999). Vaccination with cathepsin L proteinases and with leucine aminopeptidase induces high levels of protection against fascioliasis in sheep. Infect. Immun. 67, 1954–1961.
Rabut, G., and Peter, M. (2008). Function and regulation of protein neddylation. EMBO Rep. 9, 969–976. doi: 10.1038/embor.2008.183
Radulović, Ž. M., Kim, T. K., Porter, L. M., Sze, S. H., Lewis, L., and Mulenga, A. (2014). A 24-48 h fed Amblyomma americanum tick saliva immuno-proteome. BMC Genomics 15:518. doi: 10.1186/1471-2164-15-518
Ramamoorthi, N., Narasimhan, S., Pal, U., Bao, F., Yang, X. F., Fish, D., et al. (2005). The Lyme disease agent exploits a tick protein to infect the mammalian host. Nature 436, 573–577. doi: 10.1038/nature03812
Richards, S. A., Stutzer, C., Bosman, A. M., and Maritz-Olivier, C. (2015). Transmembrane proteins–mining the cattle tick transcriptome. Ticks Tick Borne Dis. 6, 695–710. doi: 10.1016/j.ttbdis.2015.06.002
Rodríguez-Mallon, A., Encinosa, P. E., Méndez-Pérez, L., Bello, Y., Fernández, R. R., Garay, H., et al. (2015). High efficacy of a 20 amino acid peptide of the acidic ribosomal protein P0 against the cattle tick, Rhipicephalus microplus. Ticks Tick Borne Dis. 6, 530–537. doi: 10.1016/j.ttbdis.2015.04.007
Rodriguez-Valle, M., Moolhuijzen, P., Barrero, R. A., Ong, C. T., Busch, G., Karbanowicz, T., et al. (2018). Transcriptome and toxin family analysis of the paralysis tick, Ixodes holocyclus. Int. J. Parasitol. 48, 71–82. doi: 10.1016/j.ijpara.2017.07.007
Sakata, E., Yamaguchi, Y., Miyauchi, Y., Iwai, K., Chiba, T., Saeki, Y., et al. (2007). Direct interactions between NEDD8 and ubiquitin E2 conjugating enzymes upregulate cullin-based E3 ligase activity. Nat. Struct. Mol. Biol. 14, 167–168. doi: 10.1038/nsmb1191
Sasaki, S. D., de Lima, C. A., Lovato, D. V., Juliano, M. A., Torquato, R. J., and Tanaka, A. S. (2008). BmSI-7, a novel subtilisin inhibitor from Boophilus microplus, with activity toward Pr1 proteases from the fungus Metarhizium anisopliae. Exp. Parasitol. 118, 214–220. doi: 10.1016/j.exppara.2007.08.003
Schulze-Lefert, P., and Panstruga, R. (2011). A molecular evolutionary concept connecting nonhost resistance, pathogen host range, and pathogen speciation. Trends Plant Sci. 16, 117–125. doi: 10.1016/j.tplants.2011.01.001
Shahein, Y. E., El-Rahim, M. T. A., Hussein, N. A., Hamed, R. R., El-Hakim, A. E., and Barakat, M. M. (2010). Molecular cloning of a small heat shock protein (sHSPII) from the cattle tick Rhipicephalus (Boophilus) annulatus salivary gland. Int. J. Biol. Macromol. 47, 614–622. doi: 10.1016/j.ijbiomac.2010.08.006
Simão, F. A., Waterhouse, R. M., Ioannidis, P., Kriventseva, E. V., and Zdobnov, E. M. (2015). BUSCO: assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 31, 3210–3212. doi: 10.1093/bioinformatics/btv351
Simiczyjew, A., Mazur, A. J., Dratkiewicz, E., and Nowak, D. (2017). Involvement of β-and γ-actin isoforms in actin cytoskeleton organization and migration abilities of bleb-forming human colon cancer cells. PLoS ONE 12:e0173709. doi: 10.1371/journal.pone.0173709
Sinha, D. K., Chandran, P., Timm, A. E., Aguirre-Rojas, L., and Smith, C. M. (2016). Virulent Diuraphis noxia aphids over-express calcium signaling proteins to overcome defenses of aphid-resistant wheat plants. PLoS ONE 11:e0146809. doi: 10.1371/journal.pone.0146809
Stark, B. C., Lanier, M. H., and Cooper, J. A. (2017). CARMIL family proteins as multidomain regulators of actin-based motility. Mol. Biol. Cell 28, 1713–1723. doi: 10.1091/mbc.e17-01-0019
Stutzer, C., van Zyl, W. A., Olivier, N. A., Richards, S., and Maritz-Olivier, C. (2013). Gene expression profiling of adult female tissues in feeding Rhipicephalus microplus cattle ticks. Int. J. Parasitol. 43, 541–554. doi: 10.1016/j.ijpara.2013.01.009
Syed, Z. A., Härd, T., Uv, A., and van Dijk-Härd, I. F. (2008). A potential role for Drosophila mucins in development and physiology. PLoS ONE 3:e3041. doi: 10.1371/journal.pone.0003041
Tan, A. W., Francischetti, I. M., Slovak, M., Kini, R. M., and Ribeiro, J. M. (2015). Sexual differences in the sialomes of the zebra tick, Rhipicephalus pulchellus. J. Proteom 117, 120–144. doi: 10.1016/j.jprot.2014.12.014
Tirloni, L., Reck, J., Terra, R. M. S., Martins, J. R., Mulenga, A., Sherman, N. E., et al. (2014). Proteomic analysis of cattle tick Rhipicephalus (Boophilus) microplus saliva: a comparison between partially and fully engorged females. PLoS ONE 9:e94831. doi: 10.1371/journal.pone.0094831
Trainor, P. A., and Merrill, A. E. (2014). Ribosome biogenesis in skeletal development and the pathogenesis of skeletal disorders. Biochim. Biophys. Acta Mol. Basis Dis. 1842, 769–778. doi: 10.1016/j.bbadis.2013.11.010
Tsubokawa, D., Hatta, T., Kikuchi, T., Maeda, H., Mikami, F., Alim, M. A., et al. (2017). Venestatin, a Ca++-binding protein from the parasitic nematode Strongyloides venezuelensis, is involved in the larval migration process. Int. J. Parasitol. 47, 501–509. doi: 10.1016/j.ijpara.2017.01.008
Turk, B. (2006). Targeting proteases: successes, failures and future prospects. Nat. Rev. Drug Discov. 5, 785–799. doi: 10.1038/nrd2092
Van Cromphaut, S. J. (2009). Hyperglycaemia as part of the stress response: the underlying mechanisms. Best Pract. Res. Clin. Anaesthesiol. 23, 375–386. doi: 10.1016/j.bpa.2009.08.005
Vedula, P., and Kashina, A. (2018). The makings of the ‘actin code’: regulation of actin's biological function at the amino acid and nucleotide level. J. Cell Sci. 131:jcs215509. doi: 10.1242/jcs.215509
Vora, A., Taank, V., Dutta, S. M., Anderson, J. F., Fish, D., Sonenshine, D. E., et al. (2017). Ticks elicit variable fibrinogenolytic activities upon feeding on hosts with different immune backgrounds. Sci. Rep. 7:44593. doi: 10.1038/srep44593
Wang, H., Zhang, X., Wang, X., Wang, M., Yang, X., Han, X., et al. (2019). Comprehensive analysis of the global protein changes that occur during salivary gland degeneration in female ixodid ticks Haemaphysalis longicornis. Front. Physiol. 9:1943. doi: 10.3389/fphys.2018.01943
Wang, J., and Maldonado, M. A. (2006). The ubiquitin-proteasome system and its role in inflammatory and autoimmune diseases. Cell Mol. Immunol. 3, 255–261.
Wang, X., Jiang, W., Kang, J., Liu, Q., and Nie, M. (2015). Knockdown of RhoA expression alters ovarian cancer biological behavior in vitro and in nude mice. Oncol. Rep. 34, 891–899. doi: 10.3892/or.2015.4009
Weisheit, S., Villar, M., Tykalová, H., Popara, M., Loecherbach, J., et al. (2015). Ixodes scapularis and Ixodes ricinus tick cell lines respond to infection with tick-borne encephalitis virus: transcriptomic and proteomic analysis. Parasit. Vectors 8:599. doi: 10.1186/s13071-015-1210-x
Williams, D. B. (2006). Beyond lectins: the calnexin/calreticulin chaperone system of the endoplasmic reticulum. J. Cell Sci. 119, 615–623. doi: 10.1242/jcs.02856
Wüthrich, M., Brandhorst, T. T., Sullivan, T. D., Filutowicz, H., Sterkel, A., Stewart, D., et al. (2015). Calnexin induces expansion of antigen-specific CD4+ T cells that confer immunity to fungal ascomycetes via conserved epitopes. Cell Host Microbe 17, 452–465. doi: 10.1016/j.chom.2015.02.009
Xu, Y., Bruno, J. F., and Luft, B. J. (2005). Identification of novel tick salivary gland proteins for vaccine development. Biochem. Biophys. Res. Commun. 326, 901–904. doi: 10.1016/j.bbrc.2004.11.127
Xu, Y., Wu, J., Fu, S., Li, C., Zhu, Z. R., and Zhou, X. (2015). Rice stripe tenuivirus nonstructural protein 3 hijacks the 26S proteasome of the small brown planthopper via direct interaction with regulatory particle non-ATPase subunit 3. J. Virol. 89, 4296–4310. doi: 10.1128/JVI.03055-14
Yang, C., Pring, M., Wear, M. A., Huang, M., Cooper, J. A., Svitkina, T. M., et al. (2005). Mammalian CARMIL inhibits actin filament capping by capping protein. Dev. Cell 9, 209–221. doi: 10.1016/j.devcel.2005.06.008
Yang, Y., Waters, J. B., Frueh, K., and Peterson, P. A. (1992). Proteasomes are regulated by interferon gamma: implications for antigen processing. Proc. Natl. Acad. Sci. U.S.A. 89, 4928–4932. doi: 10.1073/pnas.89.11.4928
You, M., and Fujisaki, K. (2009). Vaccination effects of recombinant chitinase protein from the hard tick Haemaphysalis longicornis (Acari: Ixodidae). J. Vet. Med. Sci. 71, 709–712. doi: 10.1292/jvms.71.709
Zhang, H., Forman, H. J., and Choi, J. (2005). Gamma-glutamyl transpeptidase in glutathione biosynthesis. Methods Enzymol. 401, 468–483. doi: 10.1016/S0076-6879(05)01028-1
Zhang, X., Kuramitsu, Y., Ma, A., Zhang, H., and Nakamura, K. (2016). Endoplasmic reticulium protein profiling of heat-stressed Jurkat cells by one dimensional electrophoresis and liquid chromatography tandem mass spectrometry. Cytotechnology 68, 1103–1113. doi: 10.1007/s10616-015-9867-8
Keywords: RNA-Seq, cattle tick, sialome, transcriptome, host-parasite interaction
Citation: Giachetto PF, Cunha RC, Nhani A Jr, Garcia MV, Ferro JA and Andreotti R (2020) Gene Expression in the Salivary Gland of Rhipicephalus (Boophilus) microplus Fed on Tick-Susceptible and Tick-Resistant Hosts. Front. Cell. Infect. Microbiol. 9:477. doi: 10.3389/fcimb.2019.00477
Received: 01 October 2019; Accepted: 24 December 2019;
Published: 21 January 2020.
Edited by:
Itabajara Silva Vaz Jr., Federal University of Rio Grande do Sul, BrazilReviewed by:
Iliano V. Coutinho-Abreu, National Institutes of Health (NIH), United StatesCopyright © 2020 Giachetto, Cunha, Nhani, Garcia, Ferro and Andreotti. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Renato Andreotti, cmVuYXRvLmFuZHJlb3R0aUBlbWJyYXBhLmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.