 Grith M. Maigaard Hermansen1
Grith M. Maigaard Hermansen1 Anders Boysen2
Anders Boysen2 Thøger J. Krogh2
Thøger J. Krogh2 Arkadiusz Nawrocki2
Arkadiusz Nawrocki2 Lars Jelsbak1
Lars Jelsbak1 Jakob Møller-Jensen2*
Jakob Møller-Jensen2*- 1Department of Biotechnology and Biomedicine, Technical University of Denmark, Kongens Lyngby, Denmark
- 2Department of Biochemistry and Molecular Biology, University of Southern Denmark, Odense, Denmark
Enterotoxigenic Escherichia coli (ETEC) is one of the most common causes of diarrheal illness in third world countries and it especially affects children and travelers visiting these regions. ETEC causes disease by adhering tightly to the epithelial cells in a concerted effort by adhesins, flagella, and other virulence-factors. When attached ETEC secretes toxins targeting the small intestine host-cells, which ultimately leads to osmotic diarrhea. HldE is a bifunctional protein that catalyzes the nucleotide-activated heptose precursors used in the biosynthesis of lipopolysaccharide (LPS) and in post-translational protein glycosylation. Both mechanisms have been linked to ETEC virulence: Lipopolysaccharide (LPS) is a major component of the bacterial outer membrane and is needed for transport of heat-labile toxins to the host cells, and ETEC glycoproteins have been shown to play an important role for bacterial adhesion to host epithelia. Here, we report that HldE plays an important role for ETEC virulence. Deletion of hldE resulted in markedly reduced binding to the human intestinal cells due to reduced expression of colonization factor CFA/I on the bacterial surface. Deletion of hldE also affected ETEC motility in a flagella-dependent fashion. Expression of both colonization factors and flagella was inhibited at the level of transcription. In addition, the hldE mutant displayed altered growth, increased biofilm formation and clumping in minimal growth medium. Investigation of an orthogonal LPS-deficient mutant combined with mass spectrometric analysis of protein glycosylation indicated that HldE exerts its role on ETEC virulence both through protein glycosylation and correct LPS configuration. These results place HldE as an attractive target for the development of future antimicrobial therapeutics.
Introduction
Enterotoxigenic Escherichia coli (ETEC) strains contribute significantly to diarrheal illness and mortality in third world countries (Liu et al., 2012; Platts-Mills et al., 2015). ETEC particularly affects children and is associated with millions of infections and hundreds of thousands of deaths each year but is also the most frequent cause of diarrhea among deployed military personnel and travelers visiting the endemic areas (Kotloff et al., 2013; Bourgeois et al., 2016). ETEC causes disease by adhering to epithelial cells of the upper small intestine where the delivery of heat-labile and/or heat stable toxins to host cell receptors initiates a signaling cascade, which ultimately results in watery diarrhea (Fleckenstein et al., 2013). In ETEC H10407, the host cell interaction is promoted by a combination of the plasmid-encoded fimbrial colonization factor, CFA/I, a collaboration between the adhesin EtpA and the flagellum as well as expression of virulence genes located on chromosomal pathogenicity islands (Evans et al., 1975; Patel et al., 2004; Fleckenstein et al., 2006; Roy et al., 2009b; Crossman et al., 2010). Several of the surface-exposed protein adhesins, including CFA/I and the main flagellar component FliC, have been shown to carry post-translational sugar modifications (Boysen et al., 2016). Protein glycosylation plays an important role in mediating adhesion, colonization and invasion of host tissue and may serve either as mediators of direct interactions with host-encoded cell surface glycans, as stabilizing factors of protein structure, or as a means to create surface heterogeneity and thereby evade recognition by the host immune system (Gault et al., 2015; Tytgat et al., 2016).
Lipopolysaccharide (LPS) is a central component of the Gram-negative outer membrane leaflet, forms an effective barrier against deleterious compounds, and frequently plays a role in pathogenesis (Nikaido, 2003). In ETEC, LPS is for example responsible for transport of the heat-labile toxin to host cells (Horstman and Kuehn, 2002). The LPS molecule is a three domain structure comprising (i) lipid A which anchors the LPS to the outer membrane, (ii) an inner core oligosaccharide (OS), and (iii) a O antigen polysaccharide, which is connected to the core and consists of repeating oligosaccharide units (Whitfield and Trent, 2014). The surface-exposed Lipid A and O antigen structures of an invading pathogen trigger host signaling cascades of the immune system aimed at clearing the bacterial infection (Needham and Trent, 2013). Host-driven evolution has led to numerous bacterial LPS modification strategies including glycosylation, acetylation and addition of e.g., sialic acids (Maldonado et al., 2016).
The genetics and biosynthesis pathway for lipid A and inner core OS production has been established in enteric bacteria (Frirdich and Whitfield, 2005). The lipid A synthesis pathway is highly conserved due to its role in maintaining the structural integrity of the outer membrane whereas biosynthesis of the inner core OS production displays a higher amount of structural diversity (Valvano et al., 2002). In E. coli, the inner core OS is composed of two 3-deoxy-D-manno-oct-2-ulosonic acids (Kdo) and three L-glycero-D-manno-heptose (Hep) units, which are sequentially attached to the lipid A anchor by glycosyltransferases. Mutants which are unable to synthesize the activated heptose precursor molecule or lack the enzymes required for linking of the glycans to lipid A display a characteristic phenotype referred to as “deep rough” (Frirdich and Whitfield, 2005). Collectively, this phenotype reflects changes in the outer membrane leading to changed surface hydrophilicity, which result in hypersensitivity to hydrophobic dyes, detergents, hydrophobic antibiotics, and fatty acids (Raetz and Whitfield, 2002). In E. coli, “deep rough” mutants display pleiotropic effects including bacterial auto-aggregation, loss of flagella and pili as well as elevated production of outer membrane vesicles and colanic acid exopolysaccharide (Parker et al., 1992; Nakao et al., 2012).
It has recently been determined in Shigella flexneri, Salmonella enterica serovar typhimurium, Avian pathogenic Escherichia coli (APEC), and Campylobacter jejuni that the length of LPS plays a key role in the ability of the pathogens to colonize the intestine, swarming motility, form biofilm as well as invade host cells (Kohler et al., 2002; Kong et al., 2011; Holden et al., 2012; Han et al., 2014).
Our goal of this study was to characterize the relationship between HldE and ETEC pathogenicity. HldE has an important function in the biosynthesis of ADP-activated heptose precursor units that are added to the inner core LPS (Valvano et al., 2000). In this pathway, HldE catalyzes two enzymatic steps and mutations in either of the domains resulting in truncated LPS (Kneidinger et al., 2002; Mcarthur et al., 2005). We have created an isogenic hldE mutant and report that the strain displays severely reduced adherence ability to the Caco-2 intestinal cell line consistent with an observed reduction in CFA/I expression on the cell surface. Absence of HldE also resulted in increased biofilm formation and motility defects. When using transmission electron microscopy (TEM) and western blotting, the motility defect can be linked to an absence of flagella. At the transcriptional level we show that the hldE mutation can be linked to reduced gene expression of the ETEC virulence factors FliC and colonization factor CfaB but not EtpA. In summary, our data show that HldE is needed for full virulence potential in ETEC, and that this effect is likely transmitted both through protein- and LPS heptosylation.
Materials and Methods
Bacterial Strains and Culture Conditions
Strains were grown in Luria Bertani (LB) (Sambrook and Russell, 2001) or M9 minimal medium (Clark and Maaloe, 1967) supplemented with 0.2% glucose. Cells used for electroporation were grown in Super Optimal Broth (SOB) and Super Optimal Broth with Carabolite repression (SOC) (Hanahan, 1983). Protein expression was induced from the PA1/04/03 promoter by 0.1 mM isopropyl-β-d-thiogalactopyranoside (IPTG). Ampicillin, kanamycin and chloramphenicol were supplemented when necessary. Strains and plasmids are listed in Supplementary Table S1 and primers are listed in Supplementary Table S2.
DNA Manipulations
To delete hldE and waaC in H10407 a chloramphenicol cassette was amplified from pKD3 using the primers JMJ388/JMJ389 and JMJ587/JMJ588 that entail regions flanking the hldE and waaC genes, respectively. This amplicon was introduced into H10407/pKD46 to replace the hldE and waaC genes as described by Datsenko and Wanner (2000). Plasmids transfer was carried out by electroporation (Bio-RAD gene pulser; 1.80 kV, 25 μF, 200 Ω). Electroporants were selected, isolated and tested by PCR using the primers JMJ99/JMJ391 and JMJ589/JMJ590, respectively.
Plasmids
The primer sets JMJ450/JMJ451 and JMJ589/JMJ590 were used to amplify hldE and waaC from H10407, respectively. The amplicons were digested with BamHI and XhoI and subsequently ligated into the same sites of pNDM220. The construct was verified by PCR using the primers JMJ207 and JMJ221.
Cell Line Culture Conditions
The human colon carcinoma cell line Caco-2 (Rousset, 1986) was used to study the adherence capacity. Cells were maintained in a humidified atmosphere containing 5% CO2 at 37°C and grown in Dulbecco's Modified Eagle's Medium (DMEM) (Gibco, supplied with 4.5 g/L glucose, 4.5 g/L L-glutamine and 4.5 g/L pyruvate) supplemented with 20% heat-inactivated fetal bovine serum (FBS) (Gibco) and Penicillin-Steptomycin (100 units/ml;100 μg/ml) (Gibco).
Adhesion Assay
When reaching 90% confluence the Caco-2 cells were trypsinized for 5 min, diluted 1:4 and seeded in 12-well plates (Nunc) to a density of 1∙103 cells per well. One hour prior to addition of bacteria to cell line Caco-2 cells were washed three times in PBS and incubated in DMEM without Pen-strep (Gibco). A multiplicity of infection (MOI) of 50 was used. Bacteria and cells were incubated for 2 h. Non-adherent bacteria were removed by washing the cells three times in PBS with a vigorous shake on the plate-shaker between each washing step. Relative adhesion potential was determined by serial dilutions and plating onto selective LA plates. CFU was determined next day and the adhesion ability of each strain was normalized to the adhesion ability of wild-type H10407. Results are shown as means ± standard deviations (SD).
Biofilm Formation Assay
Biofilm assays were performed in microtiter plates as described by O'Toole GA (O'toole, 2011) and Guiton et al. (2009) with a few modifications. Shortly described, ON cultures were adjusted to A600 = 0.05 before 150 μl of the diluted cultures was added to separate wells on a non-tissue culture-treated microtiter plate (Nunc). Plates were covered with microtiter sealing tape and left to incubate at 37°C for 4, 8, 24, 48 and 72 h. Planktonic bacteria were removed by submerging the plate in water and blotting excess water on a paper towel. One hundred twenty-five microliter 0.1% (w/v) crystal violet in water was added to the wells and allowed to stain for 10 min at RT. Each plate was washed 3 times as described above and left to dry ON. To solubilize the dye, 200 μl 30% acetic acid in water was added to each well. Plates were incubated with acetic acid for 10 min at RT. Acetic acid/crystal violet-solution was mixed by pipetting before 125 μl was transferred to a fresh microtiter plate. Biofilms were quantified by reading the absorbance in a VersaMax ELISA Microplate Reader (Molecular Devices) at 550 nm.
To investigate the influence of eDNA on biofilm formation 67 U/ml DNase I (Roche, RNase-free) was added to biofilm cultures from the beginning of the experiment and every 24 h thereafter. The biofilm assays were performed as described above. The biofilm formation of each strain was normalized to that of wild-type H10407. Results are shown as means ± standard deviations (SD).
Determination of Congo Red Binding
Bacterial strains were grown on LA plates and subsequently streaked on Congo red plates with and without 0.01 g/ml NaCl, respectively (0.01 g/ml tryptone, 0.005 g/ml yeast extract, 0.015 g/ml agar-agar, 40 μg/ml Congo Red, 10 μg/ml Commasie brilliant blue G-250). For plasmid-based complementation 0.1 mM IPTG was added to the plates before solidification. After incubation for 24-72 h at 28 or 37°C, respectively, plates were photographed.
Motility Assay
Swimming assay was performed in LB broth supplemented with 0.3% Difco agar and 0.4% glucose. Swarming assay was performed in LB broth supplemented with 0.45% Eiken agar and 0.4% glucose. Whenever required 0.1 mM IPTG was added to the plates. Cultures were grown in LB broth overnight and supplemented with 0.1 mM IPTG and appropriate antibiotics. The cultures were adjusted to similar densities and 2 μl were spotted in the center of the plates. The plates were then incubated at 37°C for 10 h. The diameters of motility halos were determined by using the ImageJ program (National Institute of Health). All strains were tested in biological triplicate. The halos of each strain were normalized to the halo diameter of wild-type H10407. Results are shown as means ± standard deviations (SD).
One-Dimensional SDS-PAGE and Western Blots
Culture samples were grown in LB at 37°C to exponential phase (A600 of 0.6). For detection of EtpA proteins the culture supernatants were purified by the modified Wessel-Flugge method (Wessel and Flugge, 1984). For detection of OmpA, FliC, and CfaB whole-cell lysates were used. The cell pellets were boiled in 1x SDS loading buffer (60 mM Tris-HCl, pH 6.8, 2% SDS, 10% glycerol, 0.005% bromphenol blue, 5 mM EDTA, 0.1 mM DTT) at 95°C for 5 min. The proteins were loaded onto a NUPAGE 4-12% Bis-Tris Gels (Invitrogen) for electrophoretic separation. Proteins on the gel were transferred to polyvinylidene diflouride membranes (Milipore) using transferbuffer (48 mm Tris, pH 9, 20% methanol, 39 mm glycine, 0.0375% SDS) at 0.8 mA/cm2 in a Hoefer SemiPhor blotter tank (Amersham Biosciences) for 1 h. After the transfer, the membrane was blocked with 0.3% skimmed milk in washing buffer (100 mM Tris, 150 mM NaCl, 0.05% Tween-20) and kept ON at 4°C. Alternatively immunoblotting was performed the same day. The SNAP inner diameter protein detection system 1.0 (Millipore) was used for immuno-blotting as recommended by the manufacturer. The antibodies were diluted as shown in Supplementary Table S3. Blots were developed using Western lightning reagent (PerkinElmer Life Sciences). The signal was detected and quantified using the Quantity One software associated with the ChemiDoc XRS station (Bio-Rad).
RT-qPCR
The mRNA abundance of different genes was determined by reverse transcriptase quantitative PCR (RT qPCR) using relative quantification to the ribosomal reference gene rrsA. Whole-cell lysates were harvested by growing bacteria to A600 of 0.6 and spinning at 5,000 × g for 10 min. The RNA was extracted by Hot phenol purification as previously described by Boysen et al. (2010). RNA-concentrations were determined by using a Nanodrop spectrophotometer and the integrity of the RNA was confirmed by agarose gel electrophoresis. Twenty-five microgram of RNA was DNase I-treated (Roche, RNase-free) before being reverse transcribed into cDNA (Maxima Reverse Transcriptase, Thermo Scientific) by using random hexamers. RT-qPCR samples were performed in technical duplicates in 20 μl volumes. Reactions were performed on a Stratagene MX3000P thermo cycler using the following cycling conditions: 95°C for 5 min, 40 cycles at 95°C for 15 s, 58°C for 15 s and 72°C for 15 s. Threshold cycles were analyzed using Graphpad Prism version 6.01 by two-way ANOVA analysis. The determination of the relative levels of gene expression was performed using the cycle threshold method and normalized to the reference gene rrsA. Results are represented as relative expression levels normalized to the wild-type expression level ± SD.
Transmission Electron Microscopy
For negative stain transmission electron microscopy, bacteria were grown to exponential phase in LB at 37°C. A droplet of bacterial suspension was placed on a carbon-formvar copper grid (FCF-200-Cu; Electron Microscopy Sciences, UK) for 5 min, washed three times in water and negatively stained for 30 s with 0.125% phosphotungstic acid, pH 6.0 (Sigma). A JOEL JEM-1400 electron microscope working at 120 kV was used to acquire images at 5,000x and 15,000x magnification.
Statistical Analyses
Statistical analysis was performed using Graphpad Prism version 6.01. Results are expressed as means ± SD (standard deviation). Significant differences were determined by analysis of variance using 2-way ANOVA and student's t-test. P < 0.05 was considered statistically significant.
Results
HldE Plays An Important Role in ETEC Host Cell Adherence
We constructed an ETEC H10407 strain with an isogenic hldE gene deletion and investigated its effect on ETEC adhesion to human intestinal epithelial cells. To mimic the physiological site of initial interaction, we used differentiated human intestinal Caco-2 cells with mature brush-border microvilli and tight junctions similar to what is observed in small-intestinal epithelium (Vandrangi et al., 2013). We incubated Caco-2 cells with ETEC wild type H10407/pNDM220 (wild type), the HldE-defective mutant H10407ΔhldE/pNDM220 (ΔhldE), as well as the complemented mutant H10407ΔhldE/pGH106 (ΔhldE/pGH106) with IPTG-inducible expression of the hldE gene. To validate and compare our results, a H10407ΔfliC strain was included in our experiment. The H10407ΔfliC mutant is unable to produce flagella and displays severely reduced ETEC adherence (Roy et al., 2009a). As shown in Figure 1, the adherence ability of the ΔhldE mutant to Caco-2 cells was 280-fold lower when compared to the wild-type. We also observed that complementation of the hldE gene in ΔhldE restored the binding capacity to wild type levels. In our experimental setup, the adhesion potential of the ΔhldE mutant was similar to that of the ΔfliC mutant. Taken together, these results indicate that HldE is needed for efficient adherence to Caco-2 cells either through loss of flagellae, CFA/I fimbriae or both.
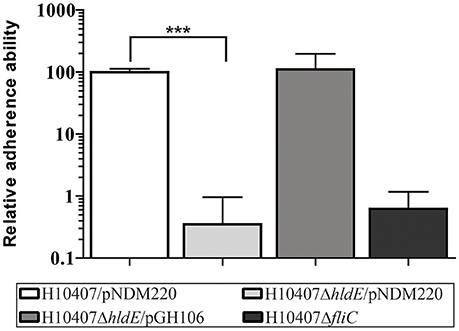
Figure 1. HldE is required for efficient host cell adhesion. The relative adhesion ability of H10407/pNDM220, H10407ΔhldE/pNDM220, H10407ΔhldE/pGH106, and H10407ΔfliC to differentiated Caco-2 cells 19 days post-seeding is shown. The adhesive capacity is relative to wild-type H10407/pNDM220 (100%). Assays were performed following induction with 0.1 mM IPTG O/N. Each time point represents three biological experiments. Values are means with standard deviations (SD). Asterisks indicate statistical significance ***P < 0.0001.
Loss of HldE in ETEC Results in Distinct Phenotypes
In E. coli K-12, deletion of the hldE gene results in a number of pronounced phenotypes including increased biofilm formation and auto-aggregation (Nakao et al., 2012). In this study we sought to characterize the phenotype of an ETEC ΔhldE mutant by evaluating biofilm formation, curli production, auto-aggregation and growth in M9 minimal medium. First, we monitored the biofilm mass produced by wild type, the HldE-defective mutant, ΔhldE, as well as the complemented mutant ΔhldE/pGH106 when grown in 96-well plates for 72 h at 37°C. The weak biofilm producing non-pathogenic E. coli strain DH5α was included in the experiment as a point of reference. The amount of formed biofilm was measured after 4, 8, 24, 48 and 72 h of static incubation. Biofilm produced by each strain was normalized to wild type levels after 4 h and plotted in Figure 2A. Over 72 h, the wild type only produced modest amounts of biofilm. Specifically, within the first 24 h all four stains produced approximately the same levels of biofilm. However, after 48 and 72 h the ΔhldE mutant had produced 20- and 30-fold more biofilm (P < 0.001), respectively, than that of the wild-type. We note that the relative increase in biofilm formation of the ΔhldE mutant is much more pronounced in ETEC when compared to an E. coli K-12 strain carrying the same genotype (Nakao et al., 2012). In this experiment, ectopic expression of the hldE gene restored biofilm formation to wild type levels (Figure 2A). It has previously been shown that the increased biofilm formation in an E. coli K-12 hldE mutant depends on the presence of extracellular DNA (Nakao et al., 2012). To investigate if the same holds true for ETEC, DNase I was added to the medium and the effect was documented. The removal of eDNA by DNase I treatment did not alter the difference in biofilm formation observed between wild-type and ΔhldE mutant (Supplementary Figure S1). In summary, the adherence to abiotic surfaces is affected by HldE in an process independent of extracellular DNA concentration, which differs from the phenotypes of the commensal E. coli K-12.
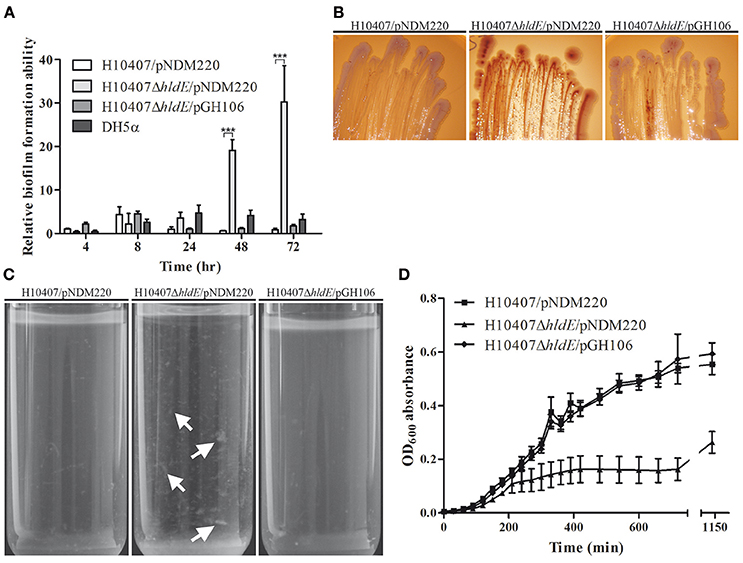
Figure 2. Phenotypic characterization of an isogenic ΔhldE deletion mutant. (A) The relative biofilm formation ability of H10407/pNDM220, H10407ΔhldE/pNDM220 and H10407ΔhldE/pGH106 is compared after 4, 8, 24, 48, and 72 h in LB media. The biofilm formation of each strain is normalized to wild-type levels at 4 h [6 biological replicates, values are means with standard deviations (SD)]. (B) The strains H10407/pNDM220, H10407ΔhldE/pNDM220 and H10407ΔhldE/pGH106 were grown on Congo Red indicator plates with 0.1 mM IPTG in order to assess the production of amyloid fibers. Representative images of strains grown at 37°C with NaCl for 72 h are shown. (C) Aggregation phenotype of ΔhldE mutant after static growth at 37°C in M9 minimal medium supplemented with 0.2% glucose. White arrows indicate aggregated cells (D) Growth of the strains H10407/pNDM220, H10407ΔhldE/pNDM220, and H10407ΔhldE/pGH106 in M9 minimal medium supplemented with 0.2% glucose at 37°C. Absorbance at OD600 was measured at the indicated time points. ***P < 0.001.
Next we analyzed the nature of the extracellular matrix formed in the biofilm assay by growing the wild type strain, ΔhldE bacteria and ΔhldE/pGH106 on Congo red indicator (CRI) plates (Figure 2B). The matrix produced by E. coli during biofilm formation mainly consists of the protein component amyloid fiber structure curli in addition to poly-β-1,6-N-acetyl-glucosamine (PGA) and/or exopolysaccharide celluloses (Danese et al., 2000; Bokranz et al., 2005; Izano et al., 2008; Smith et al., 2017). The Congo red dye will stain curli-producing colonies red whereas co-expression of both curli and cellulose results in dark purple cells. In contrast, white colonies can be observed in the absence of curli and cellulose production. Over 72 h of growth on CRI plates, a clear color difference was observed when comparing wild-type cells to ΔhldE (Figure 2B). The ΔhldE mutant colonies acquired a red color indicative of curli production exclusively, whereas the wild type cells were pale white. Complementation of the hldE isogenic deletion strain resulted in colonies with an appearance similar to that of wild type cells. Production of theses proteinaceous components also results in auto-aggregation and sedimentation of cells when grown overnight in liquid culture. To validate our observations on CRI plates, we grew wild type bacteria, the ΔhldE mutant and ΔhldE/pGH106 statically overnight in M9 minimal medium. As shown in Figure 2C, the ΔhldE mutant appeared to auto-aggregate and settle at the bottom of the test tube. In contrast, the growth medium inoculated with both the wild type and the complemented ΔhldE mutant was a homogenous suspension of bacteria.
The growth of the three ETEC strains was assessed in M9 minimal medium supplemented with 0.2% glucose (Figure 2D). In M9 minimal medium the growth of ΔhldE was significantly reduced (P < 0.0001) compared to the wild type cells. The growth defect of the mutant could be restored to wild type levels when hldE was expressed from plasmid. We note that all three strains grew similarly when cultured in LB medium (data not shown). Taken together, an isogenic hldE mutant displays enhanced biofilm formation and stain red on CRI plates and auto-aggregates in liquid culture. This indicates increased amyloid fiber production, particularly curli.
The ΔhldE Mutant is Non-Motile
It has previously been shown that deletion of hldE in E. coli K-12 results in loss of flagella (Nakao et al., 2012). To investigate if HldE is also needed for flagella production in ETEC we examined the motility ability of wild type cells, the ΔhldE mutant and ΔhldE/pGH106 as well as the non-motile ΔfliC strain using a semi-solid surface in a swimming and swarming assay. As shown in Figure 3A, the ΔhldE mutant was deficient in both swimming and swarming. To quantify the observed motility of each strain, halo diameters were measured and normalized to the wild-type cells. We observed that the ΔhldE mutant was as non-motile as the ΔfliC strain (Figures 3B,C). Moreover, plasmid based complementation of the hldE gene resulted in motility zones comparable to the wild type. We found that HldE is needed for swimming and swarming motility in ETEC.
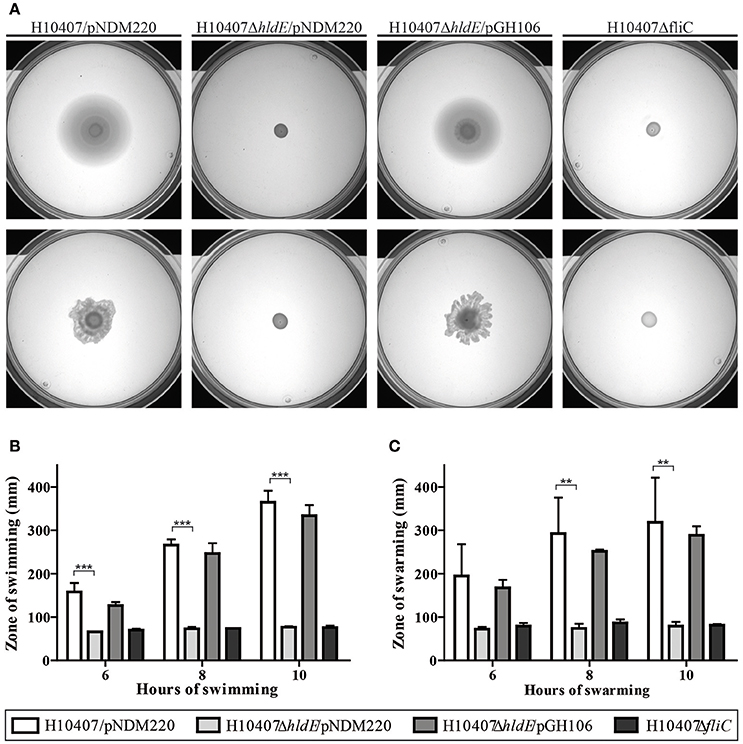
Figure 3. A ΔhldE mutant is non-motile. (A) The swimming and swarming ability of H10407/pNDM220, H10407ΔhldE/pNDM220, H10407ΔhldE/pGH106, and H10407ΔfliC is shown in top and bottom panel, respectively. (B) Quantification of the swimming halo. The swimming diameters of H10407/pNDM220, H10407ΔhldE/pNDM220, H10407ΔhldE/pGH106, and H10407ΔfliC were determined after 6, 8, and 10 h of static incubation at 37°C and plotted. (C) Quantification of the swarming halo. The swarming diameters of H10407/pNDM220, H10407ΔhldE/pNDM220, H10407ΔhldE/pGH106, and H10407ΔfliC were determined after 6, 8, and 10 h of static incubation at 37°C and plotted. Representative images are shown. All strains were tested in biological triplicate. **P < 0.01, ***P < 0.001, ns, not significant.
Aberrant Expression of Colonization Factors and Flagella in hldE Mutants
The observed motility deficiency in the ΔhldE mutant indicated an absence of flagella. In order to determine if the observed defects in cell adhesion and motility could be explained by lack of pili and flagella, respectively, we examined wild type, ΔhldE and ΔhldE/pGH106 bacteria using negative stain electron microscopy. The wild type strain appeared multi-flagellated and displayed pili on the surface (Figure 4). In contrast, the electron micrographs revealed that the ΔhldE mutant lacked flagella completely and the pili were reduced in number compared to the wild type. Moreover, the outer membrane of the mutant appeared undefined. The unusual display of surface-structures was reversed by ectopic expression of hldE from plasmid (Figure 4). In conclusion we found that deletion of hldE results in non-flagellated cells displaying altered piliation.
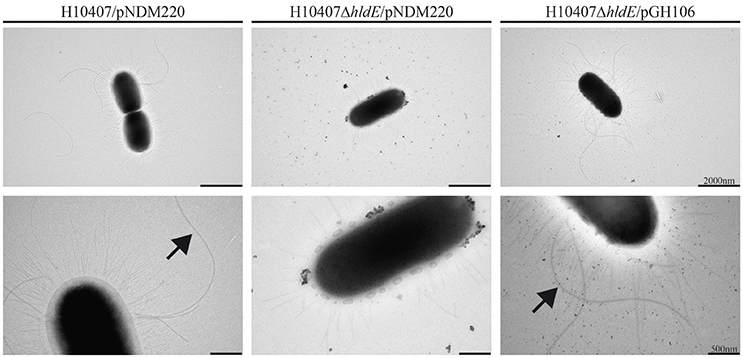
Figure 4. A ΔhldE mutant displays aberrant expression of extracellular appendages and flagella. Transmission electron micrograph analysis of the strains H10407/pNDM220, H10407ΔhldE/pNDM220, and H10407ΔhldE/pGH106. Flagella are indicated by arrowheads. Size bar is shown in the lower right corner.
HldE Affects FliC and Colonization Factor Antigen I Accumulation
The absence of flagella and aberrant pili expression in the hldE mutant strain prompted us to determine the relative levels of FliC and Colonization factor antigen I (CfaB) in whole cell lysates using Western immunoblotting. As shown in Figure 5, FliC accumulation in whole cells was significantly reduced in the ΔhldE mutant compared to the wild type and the ΔhldE mutant expressing the hldE gene from plasmid (Figure 5). To characterize the type of pili presented on the surface by the ΔhldE mutant, the level of CfaB was investigated. The western blot analysis revealed that CfaB was absent in ΔhldE mutant whole cell lysates. Plasmid based complementation restored CfaB to wild type levels (Figure 5). In summary, an isogenic ΔhldE mutant appears to affect the biosynthesis of both CfaB and flagella.
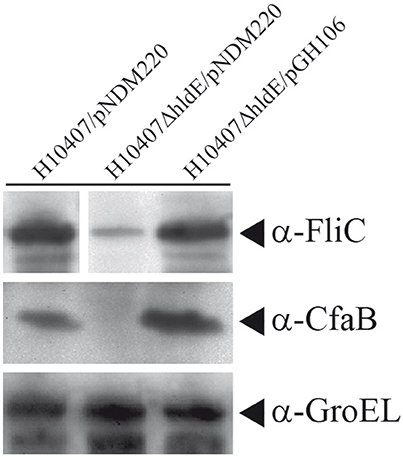
Figure 5. A ΔhldE mutant expresses reduced protein levels of the ETEC FliC and Colonization factor CfaB. Whole cell Western blot analysis of FliC and CfaB expression in strains H10407/pNDM220, H10407ΔhldE/pNDM220, and H10407ΔhldE/pGH106. GroEL chaperone protein was used as loading control. Full FliC Western blot can be viewed in Supplementary Figure S2.
HldE Affects Transcription of Flagella and Colonization Factor Antigen I
The reduced FliC and CfaB protein levels in the hldE mutant indicated decreased expression of those genes. We therefore compared the relative fliC and cfaB mRNA levels in wild type cells, the ΔhldE mutant and ΔhldE/pGH106 bacteria using RT-qPCR (Figures 6A,B). As depicted in Figure 6A, the relative fliC mRNA expression level in the ΔhldE mutant was ~15 to 20-fold reduced compared to wild type strain. Again, complementation with hldE from plasmid restored the fliC expression to wild type levels.
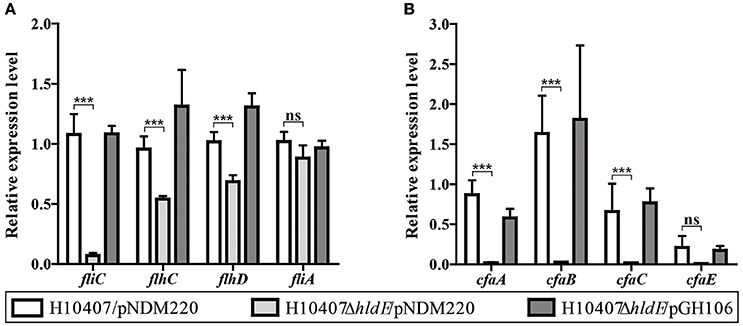
Figure 6. HldE is required for appropriate expression of genes involved in the synthesis of flagella and colonization factor antigen I. RT-qPCR was used to analyze the relative fliC, flhCD, fliA, and cfa operon mRNA transcript levels in strains H10407/pNDM220, H10407ΔhldE/pNDM220, and H10407ΔhldE/pGH106. (A) Gene expression ratios of fliC as well as Class I and Class II genes, flhCD, and fliA, respectively, involved in the flagella synthesis is shown. (B) Gene expression profile of the cfa operon genes, cfaA, cfaB cfaC, and cfaE is depicted. Bars represent mean fold changes and standard deviations are indicated. The results are based on two biological experiments. The data is normalized to wild-type levels in one experiment. The gene rrsA was used as internal reference for normalization. The standard deviations are indicated. ***P < 0.001, ns, not significant.
Flagellar synthesis is highly regulated and depends on the Class I master regulators flhDC as well as the Class II alternative sigma factor fliA (Fitzgerald et al., 2014). Next, we sought to determine the relative Class I and Class II mRNA levels in the three strains. As presented in Figure 6A, we observed a significant decrease of the flhD and flhC mRNA expression levels in the ΔhldE mutant compared to wild type and ΔhldE/pGH106. These results indicate that the deletion of hldE not only affects fliC but also Class I genes.
Similarly, using RT-qPCR we examined the relative mRNA levels of cfaB in wild type cells and ΔhldE mutant. The cfaA-E operon is a four-gene polycistronic mRNA of which cfaB is the second product (Jordi et al., 1992b). As shown in Figure 6B, the mRNA transcripts encoding cfaA and cfaB were 50- and 65-fold more abundant in both wild-type and ΔhldE/pGH106 compared to the ΔhldE mutant. The relative difference of cfaCE mRNA levels between wild type and ΔhldE mutant were lower but still 30- and 20-fold reduced, respectively. We note, that although the cfa operon is transcribed as a polycistronic messenger under the control of a single distant promoter, higher levels of cfaB accumulate in the cell compared to cfaA and cfaCE (Figure 6B). This pattern of differential stability has previously been reported (Jordi et al., 1993).
Taken together, the RT-qPCR results show that HldE is needed for proper expression of both flagella synthesis genes as well as the entire cfa operon.
HldE Does Not Affect Transcription of EtpA and OmpA
In addition to the flagellum and Colonization factor antigen I, ETEC employs a panel of surface located proteins which all contribute to the host cell interaction. One of these proteins is the glycosylated adhesin EtpA (Fleckenstein et al., 2006). We asked whether the absence of HldE would influence the expression level of the adhesin EtpA. Using RT-qPCR, we compared the relative etpA mRNA levels in wild type, ΔhldE mutant and bacteria (Figure 7A). As depicted in Figure 7A, the etpA mRNA levels were comparable in all three strains. At the protein level, the EtpA accumulation profile was similar in all three strains (Figure 7B).
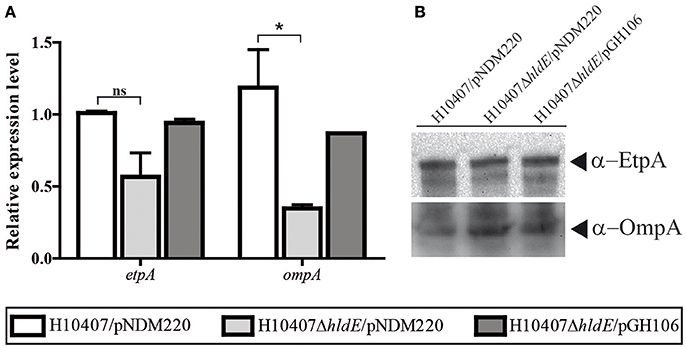
Figure 7. The relative mRNA and protein levels of ETEC virulence factor etpA and outer membrane protein A, ompA, was analyzed in the strains H10407/pNDM220, H10407ΔhldE/pNDM220, and H10407ΔhldE/pGH106. (A) The relative etpA and ompA mRNA expression levels was determined using RT-qPCR. The results are based on two biological experiments performed in technical triplicate. The data is normalized to wild-type levels in one experiment. The gene rrsA was used as internal reference for normalization. The standard deviations are indicated. *P < 0.05, ns, not significant. (B) The EtpA and OmpA protein accumulation profile was determined by Western blot analysis.
The outer membrane protein OmpA is not just one of the most abundant molecules synthesized in E. coli but it is also crucial for host cell receptor binding in a number of pathogenic invasive E. coli strains (Sugawara and Nikaido, 1994; Prasadarao, 2002; Rolhion et al., 2010). We investigated the OmpA expression at both the protein and transcriptional level in order to assign a potential role in ETEC pathogenesis. As shown in Figure 7A, we measured reduced ompA mRNA levels (P < 0.05) in a ΔhldE mutant compared to wild type bacteria. In this experiment, induction of hldE from plasmid restored mRNA expression to wild type levels (Figure 7A). On the other hand, in whole cell lysate samples, the OmpA levels in the mutant were comparable to the wild type and ΔhldE/pGH106 (Figure 7B). In conclusion, an isogenic ΔhldE mutant does not affect expression of EtpA. Reduced transcription of ompA was observed but the effect was not evident on the protein level.
HldE Deletion Does Not Abolish Protein Glycosylation
Due to the dual function of HldE in LPS biosynthesis and protein glycosylation, the reduced virulence potential observed in the ΔhldE mutant could have two explanations. In attempt to discriminate between LPS-mediated and glycoprotein-mediated effects, we analyzed an orthogonal LPS-defective mutant strain H10407ΔwaaC, which lacked heptosyl transferase I activity required for LPS core biosynthesis (Nakao et al., 2012). Motility assays confirmed that H10407ΔwaaC was severely impaired in both swimming and swarming motility at 10 h post-incubation (Figures 8A,B). Western blot analysis confirmed that FliC expression was reduced similar to what was observed for H10407ΔhldE (Figure 8D). However, there was no significant reduction in the ability of H10407ΔwaaC to adhere to differentiated Caco-2 cells (Figure 8C), indicating that the adhesion defect displayed by HldE deficient ETEC is not entirely LPS dependent. We next determined the glycoprotein profile of wild type H10407 and its isogenic ΔhldE mutant by BEMAP analysis (Boysen et al., 2016). As shown in Supplementary Table S4, the glycopeptide profile was-with some notable exceptions-largely unaffected by deletion of hldE, indicating the ETEC employs other types of sugar modifications in addition to heptose on its surface. Consistent with the lack of flagellar gene transcription, FliC glycopeptides were absent in the HldE deficient mutant. The detection of CfaB glycopeptides in both strains despite the significant reduction in pili biosynthesis in the ΔhldE mutant is not surprising given that the BEMAP technique allows for enrichment of glycopeptides from a complex sample. Notably, the flu gene product Antigen 43-another major protein adhesion-was found to be absent in the ΔhldE mutant.
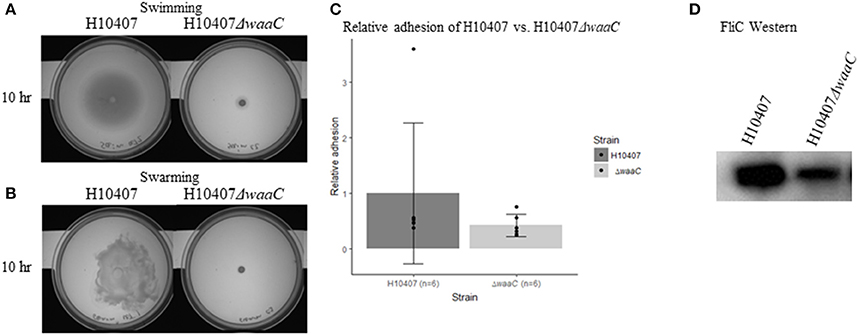
Figure 8. LPS-defective H10407ΔwaaC phenotypes. (A) Swimming- and (B) swarming motility of H10407 wild type and H10407ΔwaaC after 10 h. (C) Relative adhesion of H10407 and H10407ΔwaaC to differentiated Caco-2 cells 19 days post-seeding. The adhesive capacity is relative to the average adhesion of wild-type H10407. (D) Western blot showing FliC levels in wild type H10407 and H10407ΔwaaC cells.
Discussion
The ability of enteric pathogens to colonize the human intestine is the first step in an orchestrated host cell engagement. In S. flexneri and S. enterica serovar typhimurium both LPS and protein based virulence factors play roles in the initial attachment (Kohler et al., 2002; Yoon et al., 2009; Kong et al., 2011; Mattock and Blocker, 2017). As HldE has been reported to play a role in both LPS biosynthesis and protein glycosylation, we wanted to investigate the potential link between HldE and virulence in ETEC. This was attempted by generating an isogenic knockout mutant in the hldE gene, which plays a role in the biosynthesis of ADP-activated heptose units required for a structurally complete LPS (Valvano et al., 2000) as well as for post-translational protein heptosylation.
We have shown that an hldE mutant was severely impaired in adhesion to differentiated and polarized Caco-2 cells when compared to wild type bacteria and a non-motile ΔfliC strain (Figure 1). Moreover, in examining the ΔhldE strain using TEM and motility assays, we observed that the mutant was non-motile (Figure 3) and displayed aberrant expression of colonization factor and flagella on the surface of the bacteria (Figure 4). For C. jejuni and S. enterica serovar typhimurium it has previously been reported that functional LPS is needed for bacterial motility (Toguchi et al., 2000; Holden et al., 2012). Our results confirm that this is also the case for ETEC. Consistently, an orthogonal LPS-defective mutant H10407ΔwaaC, which was non-motile at 10 h post-inoculation in swimming and swarming motility assays, similar to the ΔhldE mutant (Figure 8). However, the ΔwaaC mutant displayed a much weaker reduction in cell adhesion compared to ΔhldE, indicating that defect LPS cannot fully account for all the observed hldE phenotypes. Thus, HldE is required for optimal cell adherence via a non-LPS-dependent mechanism, which likely involved glycoprotein modification.
It has recently been shown that ETEC extensively O-glycosylates its proteome (Boysen et al., 2016). The list of identified glycosylated proteins includes metabolic enzymes, outer membrane transporters and the majority of bona fide virulence factors identified in H10407 such as Flagellin, Ag43, EtpA, EatA, TibA, CfaB, CexE and an auto-transporter sharing homology with Ag43. The glycoproteins TibA and Ag43 are modified with heptose monosaccharides whereas EtpA potentially carries N-acetylglucosamine (GlcNAc) glycans (Lindenthal and Elsinghorst, 1999; Benz and Schmidt, 2001; Fleckenstein et al., 2006; Sherlock et al., 2006). The glycans used for protein glycosylation is redirected from the biosynthesis pathway producing the activated heptose precursor molecules intended for LPS production (Benz and Schmidt, 2001). It has been shown that protein glycosylation influences stability, functionality, and the host cell adhesive ability (Lindenthal and Elsinghorst, 2001; Fleckenstein et al., 2006; Sherlock et al., 2006; Knudsen et al., 2008; Cote et al., 2013). Our hldE mutant is unable to synthesize the heptose precursor molecules for both the LPS core OS production and protein glycosylation. We speculate that the reduced Caco-2 cell line adherence ability of the mutant could be a cumulative effect of both non-functional virulence factors and truncated LPS.
Western blotting further demonstrated that the ΔhldE mutant failed to accumulate both FliC and colonization factor CFA/I (Figure 5). Using RT-qPCR, we discovered that the gene deletion significantly reduced the mRNA transcript levels of the regulatory Class I genes flhCD and the fliC gene (Figure 6A). Furthermore, the cfaABC mRNA levels of the CFA/I operon were affected in ΔhldE when compared to wild-type (Figure 6B). From the expression analysis, it became evident that the pili expressed on the surface of hldE mutants as seen in Figure 4 are likely not CFA/I. Instead, our BEMAP analysis selectively identified glycopeptides from Type 1 fimbrial outer membrane usher proteins in the ΔhldE mutant. It seems plausible that the pili displayed by the mutant entail a different tissue tropism possibly explaining the reduced adherence capacity (Figure 1). We also examined the relative gene expression levels of the adhesin EtpA and the outer membrane porin OmpA. As shown in Figure 7, deleting hldE did not significantly affect the etpA mRNA levels or protein abundance when comparing to wild type cells. In contrast, we observed reduced amounts of ompA mRNA, which was not evident at the protein level.
At the mRNA level, fliC expression is downregulated by the σE response in Salmonella (Li et al., 2015) and we detect a similar decrease in fliC mRNA in our ETEC hldE mutant. On the other hand, we observed a downregulation of the Class I flhCD regulators in the ΔhldE strain, which does not occur in Salmonella (Li et al., 2015). Moreover, a Salmonella mutant carrying a LPS structure similar to our hldE strain produces wild type levels of FliC contrary to our observations (Toguchi et al., 2000). Taken together, by comparing the σE response in Salmonella to our data, it is plausible that the ETEC σE network conveys input to the flagellum operon and the ompA gene, which results in different outcomes. Further analyses are required in order to unravel the LPS feedback and potential σE-regulated induction of virulence factors in a ΔhldE ETEC strain.
Expression of the ETEC H10407 colonization factor CFA/I and the virulence factor EtpA has been studied in great detail. The cfaABCE operon is positively regulated by the trans-encoded CfaD protein belonging to the family of AraC transcriptional regulators and negatively regulated by histone-like protein H-NS and cAMP receptor protein (CRP) (Caron and Scott, 1990; Jordi et al., 1992a; Munson and Scott, 1999; Pilonieta et al., 2007; Bodero and Munson, 2009). In addition, several environmental factors of the intestinal lumen influence CFA/I expression including iron levels, pH, and gastric mucin (Haines et al., 2015). The etpBAC and cfaABCE operons are located on the same plasmid carried by ETEC and share inducing cues such as iron starvation. Our results demonstrate a link between a truncated LPS structure and cfa gene expression but not to etpA. In summary, we have shown that HldE activity is needed for proper expression of the CFA/I colonization factor as deletion of hldE leads to reduced transcription of the cfaABCE operon and reduced protein levels. Whether this effect is mediated by the the σE response in response to envelope stress caused by LPS alteration remains to be determined. Alternatiely, changes in gene expression could result from a lack of transcription factor activation through glycosylation, as described for eukaryotic transcriptions factors (Jackson and Tjian, 1988). Indeed, an uncharacterised LysR-family transcription factor (ETEC1629) was found to be non-glycosylated in the hldE deletion mutant (Supplementary Table S4).
By truncating the LPS structure in ETEC we observed phenotypes that also have been described in Avian Pathogenic E. coli (APEC) and non-pathogenic E. coli K12 mutants (Nakao et al., 2012; Han et al., 2014). The ΔhldE strain produced significantly more biofilm after 48 and 72 h compared to wild type cells (Figure 2A). However, the mutant biofilm was different from extracellular matrix produced by other bacteria. Both Listeria monocytogenes and Pseudomonas aeruginosa produce substantial quantities of eDNA, which stabilize the biofilm structures (Whitchurch et al., 2002; Harmsen et al., 2010). These biofilms are sensitive to DNase I treatment just as the structures produced by E. coli K-12 LPS mutants (Nakao et al., 2012). In our experiments, DNase I treatment did not significantly change the amount of biofilm formation in the mutant background (Supplemental Figure S1). This indicates that the biofilm was primarily composed of proteinaceous components. Based on CRI plates, the hldE mutant biofilm appeared to contain curli (Figure 2B). The fact that curli are not evident on the electron micrograph might reflect differences in growth conditions of the two assays as cells investigated by TEM where grown in suspension. Another property of biofilm forming bacteria is the ability to auto-aggregate and settle at the bottom of a test tube when grown statically. The capability to sediment has been shown in E. coli K12, diarrhea-causing E. coli, UPEC and ETEC to depend on the glycosylated cell surface-displayed autotransporters Ag43, AidA, and TibA as well as pili (Sherlock et al., 2004, 2005, 2006; Liaqat and Sakellaris, 2012; Cote et al., 2013). Using TEM, we observed that the hldE mutant presented abnormal pili on the surface, which indicates that this extracellular appendage is unlikely to promote the increased biofilm formation. The proteins Ag43 and TibA are post-translationally modified with heptose glycans, which the ΔhldE strain is unable to synthesize (Benz and Schmidt, 2001; Sherlock et al., 2005, 2006). We do however note that protein glycosylation is not required for promoting auto-aggregation. Taken together, the biofilm formed by the mutant cells is likely to consist of both curli and surface exposed autotransporters.
The importance of LPS modifications in determining resistance to polymyxins and bacterial infection establishment has been demonstrated in multiple bacterial pathogens (Beceiro et al., 2014; Olaitan et al., 2014). Our data groups ETEC with pathogens such as S. flexneri, S. enterica serovar typhimurium, and C. jejuni in which the length of LPS plays a key role in the ability to colonize the intestine (Kohler et al., 2002; Kong et al., 2011; Holden et al., 2012). Moreover, given the abundance of protein glycosylation, we are investigating the glycoproteome of wild type bacteria and an hldE mutant in order to determine which proteins are heptosylated and if they possess particular properties. These studies will reveal which factors of the hldE mutant phenotypes that involve altered protein glycosylation. Knowledge about the impact of truncated LPS on ETEC virulence will be valuable for vaccine development and the data presented here reveal that HldE could be a potential future therapeutic target as its deletion results in reduced bacterial virulence.
Author Contributions
Studies were designed by GM, AB, TK, JM-J. Experiments were conducted by GM, TK, AB, and AN. Data interpretation was performed by GM, AB, and JM-J. GM worked under the supervision of LJ and JM-J. Manuscript was written by GM and AB and revised, approved by all authors.
Funding
LJ acknowledges funding from the Villum Foundation (grant number VKR023113). AB acknowledges support from the Lundbeck Foundation (grant number R31-A2459) and The Villum Foundation (grant number 436879).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2018.00253/full#supplementary-material
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Beceiro, A., Moreno, A., Fernández, N., Vallejo, J. A., Aranda, J., Adler, B., et al. (2014). Biological cost of different mechanisms of colistin resistance and their impact on virulence in Acinetobacter baumannii. Antimicrob. Agents Chemother. 58, 518–526. doi: 10.1128/AAC.01597-13
Benz, I., and Schmidt, M. A. (2001). Glycosylation with heptose residues mediated by the aah gene product is essential for adherence of the AIDA-I adhesin. Mol. Microbiol. 40, 1403–1413. doi: 10.1046/j.1365-2958.2001.02487.x
Bodero, M. D., and Munson, G. P. (2009). Cyclic AMP receptor protein-dependent repression of heat-labile enterotoxin. Infect. Immun. 77, 791–798. doi: 10.1128/IAI.00928-08
Bokranz, W., Wang, X., Tschäpe, H., and Römling, U. (2005). Expression of cellulose and curli fimbriae by Escherichia coli isolated from the gastrointestinal tract. J. Med. Microbiol. 54, 1171–1182. doi: 10.1099/jmm.0.46064-0
Bourgeois, A. L., Wierzba, T. F., and Walker, R. I. (2016). Status of vaccine research and development for enterotoxigenic Escherichia coli. Vaccine 34, 2880–2886. doi: 10.1016/j.vaccine.2016.02.076
Boysen, A., Møller-Jensen, J., Kallipolitis, B., Valentin-Hansen, P., and Overgaard, M. (2010). Translational regulation of gene expression by an anaerobically induced small non-coding RNA in Escherichia coli. J. Biol. Chem. 285, 10690–10702. doi: 10.1074/jbc.M109.089755
Boysen, A., Palmisano, G., Krogh, T. J., Duggin, I. G., Larsen, M. R., and Moller-Jensen, J. (2016). A novel mass spectrometric strategy “BEMAP” reveals Extensive O-linked protein glycosylation in Enterotoxigenic Escherichia coli. Sci. Rep. 6:32016. doi: 10.1038/srep32016
Caron, J., and Scott, J. R. (1990). A rns-like regulatory gene for colonization factor antigen I (CFA/I) that controls expression of CFA/I pilin. Infect. Immun. 58, 874–878.
Clark, J. D., and Maaloe, O. (1967). DNA replication and the division cycle in Escherichia coli. J. Mol. Biol. 23, 99–112. doi: 10.1016/S0022-2836(67)80070-6
Côté, J. P., Charbonneau, M. É., and Mourez, M. (2013). Glycosylation of the Escherichia coli TibA self-associating autotransporter influences the conformation and the functionality of the protein. PLoS ONE 8:e80739. doi: 10.1371/journal.pone.0080739
Crossman, L. C., Chaudhuri, R. R., Beatson, S. A., Wells, T. J., Desvaux, M., Cunningham, A. F., et al. (2010). A commensal gone bad: complete genome sequence of the prototypical enterotoxigenic Escherichia coli strain H10407. J. Bacteriol. 192, 5822–5831. doi: 10.1128/JB.00710-10
Danese, P. N., Pratt, L. A., and Kolter, R. (2000). Exopolysaccharide production is required for development of Escherichia coli K-12 biofilm architecture. J. Bacteriol. 182, 3593–3596. doi: 10.1128/JB.182.12.3593-3596.2000
Datsenko, K. A., and Wanner, B. L. (2000). One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U.S.A. 97, 6640–6645. doi: 10.1073/pnas.120163297
Evans, D. G., Silver, R. P., Evans, D. J., Chase, D. G., and Gorbach, S. L. (1975). Plasmid-controlled colonization factor associated with virulence in Esherichia coli enterotoxigenic for humans. Infect. Immun. 12, 656–667.
Fitzgerald, D. M., Bonocora, R. P., and Wade, J. T. (2014). Comprehensive mapping of the Escherichia coli flagellar regulatory network. PLoS Genet. 10:e1004649. doi: 10.1371/journal.pgen.1004649
Fleckenstein, J. M., Munson, G. M., and Rasko, D. (2013). Enterotoxigenic Escherichia coli: Orchestrated host engagement. Gut Microb. 4, 392–396. doi: 10.4161/gmic.25861
Fleckenstein, J. M., Roy, K., Fischer, J. F., and Burkitt, M. (2006). Identification of a two-partner secretion locus of enterotoxigenic Escherichia coli. Infect. Immun. 74, 2245–2258. doi: 10.1128/IAI.74.4.2245-2258.2006
Frirdich, E., and Whitfield, C. (2005). Lipopolysaccharide inner core oligosaccharide structure and outer membrane stability in human pathogens belonging to the Enterobacteriaceae. J. Endotoxin Res. 11, 133–144. doi: 10.1179/096805105X46592
Gault, J., Ferber, M., Machata, S., Imhaus, A. F., Malosse, C., Charles-Orszag, A., et al. (2015). Neisseria meningitidis Type IV Pili composed of sequence invariable pilins are masked by multisite glycosylation. PLoS Pathog. 11:e1005162. doi: 10.1371/journal.ppat.1005162
Guiton, P. S., Hung, C. S., Kline, K. A., Roth, R., Kau, A. L., Hayes, E., et al. (2009). Contribution of autolysin and Sortase a during Enterococcus faecalis DNA-dependent biofilm development. Infect. Immun. 77, 3626–3638. doi: 10.1128/IAI.00219-09
Haines, S., Gautheron, S., Nasser, W., and Renauld-Mongénie, G. (2015). Identification of novel components influencing colonization factor antigen I expression in enterotoxigenic Escherichia coli. PLoS ONE 10, e0141469. doi: 10.1371/journal.pone.0141469
Han, Y., Han, X., Wang, S., Meng, Q., Zhang, Y., Ding, C., et al. (2014). The waaL gene is involved in lipopolysaccharide synthesis and plays a role on the bacterial pathogenesis of avian pathogenic Escherichia coli. Vet. Microbiol. 172, 486–491. doi: 10.1016/j.vetmic.2014.05.029
Hanahan, D. (1983). Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 166, 557–580. doi: 10.1016/S0022-2836(83)80284-8
Harmsen, M., Lappann, M., Knøchel, S., and Molin, S. (2010). Role of extracellular DNA during biofilm formation by Listeria monocytogenes. Appl. Environ. Microbiol. 76, 2271–2279. doi: 10.1128/AEM.02361-09
Holden, K. M., Gilbert, M., Coloe, P. J., Li, J., and Fry, B. N. (2012). The role of WlaRG, WlaTB and WlaTC in lipooligosaccharide synthesis by Campylobacter jejuni strain 81116. Microb. Pathog. 52, 344–352. doi: 10.1016/j.micpath.2012.03.004
Horstman, A. L., and Kuehn, M. J. (2002). Bacterial surface association of heat-labile enterotoxin through lipopolysaccharide after secretion via the general secretory pathway. J. Biol. Chem. 277, 32538–32545. doi: 10.1074/jbc.M203740200
Izano, E. A., Sadovskaya, I., Wang, H., Vinogradov, E., Ragunath, C., Ramasubbu, N., et al. (2008). Poly-N-acetylglucosamine mediates biofilm formation and detergent resistance in Aggregatibacter actinomycetemcomitans. Microb. Pathog. 44, 52–60. doi: 10.1016/j.micpath.2007.08.004
Jackson, S. P., and Tjian, R. (1988). O-glycosylation of eukaryotic transcription factors: implications for mechanisms of transcriptional regulation. Cell 55, 125–133. doi: 10.1016/0092-8674(88)90015-3
Jordi, B. J., Dagberg, B., de Haan, L. A., Hamers, A. M., Van der Zeijst, B. A., Gaastra, W., et al. (1992a). The positive regulator CfaD overcomes the repression mediated by histone-like protein H-NS (H1) in the CFA/I fimbrial operon of Escherichia coli. EMBO J. 11, 2627–2632.
Jordi, B. J., Willshaw, G. A., Van der Zeijst, B. A., and Gaastra, W. (1992b). The complete nucleotide sequence of region 1 of the CFA/I fimbrial operon of human enterotoxigenic Escherichia coli. DNA Seq. 2, 257–263. doi: 10.3109/10425179209020811
Jordi, B. J., Op den Camp, I. E., de Haan, L. A., Van der Zeijst, B. A., and Gaastra, W. (1993). Differential decay of RNA of the CFA/I fimbrial operon and control of relative gene expression. J. Bacteriol. 175, 7976–7981. doi: 10.1128/jb.175.24.7976-7981.1993
Kneidinger, B., Marolda, C., Graninger, M., Zamyatina, A., Mcarthur, F., Kosma, P., et al. (2002). Biosynthesis pathway of ADP-L-glycero-beta-D-manno-heptose in Escherichia coli. J. Bacteriol. 184, 363–369. doi: 10.1128/JB.184.2.363-369.2002
Knudsen, S. K., Stensballe, A., Franzmann, M., Westergaard, U. B., and Otzen, D. E. (2008). Effect of glycosylation on the extracellular domain of the Ag43 bacterial autotransporter: enhanced stability and reduced cellular aggregation. Biochem. J. 412, 563–577. doi: 10.1042/BJ20071497
Köhler, H., Rodrigues, S. P., and Mccormick, B. A. (2002). Shigella flexneri interactions with the basolateral membrane domain of polarized model intestinal epithelium: role of lipopolysaccharide in cell invasion and in activation of the mitogen-activated protein kinase ERK. Infect. Immun. 70, 1150–1158. doi: 10.1128/IAI.70.3.1150-1158.2002
Kong, Q., Yang, J., Liu, Q., Alamuri, P., Roland, K. L., and Curtiss, R. III. (2011). Effect of deletion of genes involved in lipopolysaccharide core and O-antigen synthesis on virulence and immunogenicity of Salmonella enterica serovar typhimurium. Infect. Immun. 79, 4227–4239. doi: 10.1128/IAI.05398-11
Kotloff, K. L., Nataro, J. P., Blackwelder, W. C., Nasrin, D., Farag, T. H., Panchalingam, S., et al. (2013). Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet 382, 209–222. doi: 10.1016/S0140-6736(13)60844-2
Li, J., Overall, C. C., Nakayasu, E. S., Kidwai, A. S., Jones, M. B., Johnson, R. C., et al. (2015). Analysis of the Salmonella regulatory network suggests involvement of SsrB and H-NS in sigma(E)-regulated SPI-2 gene expression. Front. Microbiol. 6, 27. doi: 10.3389/fmicb.2015.00027
Liaqat, I., and Sakellaris, H. (2012). Biofilm formation and binding specificities of CFA/I, CFA/II and CS2 adhesions of enterotoxigenic Escherichia coli and Cfae-R181A mutant. Braz. J. Microbiol. 43, 969–980. doi: 10.1590/S1517-83822012000300018
Lindenthal, C., and Elsinghorst, E. A. (1999). Identification of a glycoprotein produced by enterotoxigenic Escherichia coli. Infect. Immun 67, 4084–4091.
Lindenthal, C., and Elsinghorst, E. A. (2001). Enterotoxigenic Escherichia coli TibA glycoprotein adheres to human intestine epithelial cells. Infect. Immun. 69, 52–57. doi: 10.1128/IAI.69.1.52-57.2001
Liu, L., Johnson, H. L., Cousens, S., Perin, J., Scott, S., Lawn, J. E., et al. (2012). Global, regional, and national causes of child mortality: an updated systematic analysis for 2010 with time trends since 2000. Lancet 379, 2151–2161. doi: 10.1016/S0140-6736(12)60560-1
Maldonado, R. F., Sá-Correia, I., and Valvano, M. A. (2016). Lipopolysaccharide modification in Gram-negative bacteria during chronic infection. FEMS Microbiol. Rev. 40, 480–493. doi: 10.1093/femsre/fuw007
Mattock, E., and Blocker, A. J. (2017). How do the virulence factors of Shigella work together to cause disease? Front. Cell. Infect. Microbiol. 7, 64. doi: 10.3389/fcimb.2017.00064
Mcarthur, F., Andersson, C. E., Loutet, S., Mowbray, S. L., and Valvano, M. A. (2005). Functional analysis of the glycero-manno-heptose 7-phosphate kinase domain from the bifunctional HldE protein, which is involved in ADP-L-glycero-D-manno-heptose biosynthesis. J. Bacteriol. 187, 5292–5300. doi: 10.1128/JB.187.15.5292-5300.2005
Munson, G. P., and Scott, J. R. (1999). Binding site recognition by Rns, a virulence regulator in the AraC family. J. Bacteriol. 181, 2110–2117.
Nakao, R., Ramstedt, M., Wai, S. N., and Uhlin, B. E. (2012). Enhanced biofilm formation by Escherichia coli LPS mutants defective in Hep biosynthesis. PLoS ONE 7:e51241. doi: 10.1371/journal.pone.0051241
Needham, B. D., and Trent, M. S. (2013). Fortifying the barrier: the impact of lipid A remodelling on bacterial pathogenesis. Nat. Rev. Microbiol. 11, 467–481. doi: 10.1038/nrmicro3047
Nikaido, H. (2003). Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 67, 593–656. doi: 10.1128/MMBR.67.4.593-656.2003
Olaitan, A. O., Morand, S., and Rolain, J. M. (2014). Mechanisms of polymyxin resistance: acquired and intrinsic resistance in bacteria. Front. Microbiol. 5, 643. doi: 10.3389/fmicb.2014.00643
O'Toole, G. A. (2011). Microtiter dish biofilm formation assay. J. Vis. Exp. 47, 2437. doi: 10.3791/2437
Parker, C. T., Kloser, A. W., Schnaitman, C. A., Stein, M. A., Gottesman, S., and Gibson, B. W. (1992). Role of the rfaG and rfaP genes in determining the lipopolysaccharide core structure and cell surface properties of Escherichia coli K-12. J. Bacteriol. 174, 2525–2538. doi: 10.1128/jb.174.8.2525-2538.1992
Patel, S. K., Dotson, J., Allen, K. P., and Fleckenstein, J. M. (2004). Identification and molecular characterization of EatA, an autotransporter protein of enterotoxigenic Escherichia coli. Infect. Immun. 72, 1786–1794. doi: 10.1128/IAI.72.3.1786-1794.2004
Pilonieta, M. C., Bodero, M. D., and Munson, G. P. (2007). CfaD-dependent expression of a novel extracytoplasmic protein from enterotoxigenic Escherichia coli. J. Bacteriol. 189, 5060–5067. doi: 10.1128/JB.00131-07
Platts-Mills, J. A., Babji, S., Bodhidatta, L., Gratz, J., Haque, R., Havt, A., et al. (2015). Pathogen-specific burdens of community diarrhoea in developing countries: a multisite birth cohort study (MAL-ED). Lancet Glob. Health 3, e564–e575. doi: 10.1016/S2214-109X(15)00151-5
Prasadarao, N. V. (2002). Identification of Escherichia coli outer membrane protein A receptor on human brain microvascular endothelial cells. Infect. Immun. 70, 4556–4563. doi: 10.1128/IAI.70.8.4556-4563.2002
Raetz, C. R., and Whitfield, C. (2002). Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 71, 635–700. doi: 10.1146/annurev.biochem.71.110601.135414
Rolhion, N., Barnich, N., Bringer, M. A., Glasser, A. L., Ranc, J., Hébuterne, X., et al. (2010). Abnormally expressed ER stress response chaperone Gp96 in CD favours adherent-invasive Escherichia coli invasion. Gut 59, 1355–1362. doi: 10.1136/gut.2010.207456
Rousset, M. (1986). The human colon carcinoma cell lines HT-29 and Caco-2: two in vitro models for the study of intestinal differentiation. Biochimie 68, 1035–1040. doi: 10.1016/S0300-9084(86)80177-8
Roy, K., Hamilton, D., Ostmann, M. M., and Fleckenstein, J. M. (2009a). Vaccination with EtpA glycoprotein or flagellin protects against colonization with enterotoxigenic Escherichia coli in a murine model. Vaccine 27, 4601–4608. doi: 10.1016/j.vaccine.2009.05.076
Roy, K., Hilliard, G. M., Hamilton, D. J., Luo, J., Ostmann, M. M., and Fleckenstein, J. M. (2009b). Enterotoxigenic Escherichia coli EtpA mediates adhesion between flagella and host cells. Nature 457, 594–598. doi: 10.1038/nature07568
Sambrook, J., and Russell, D. (2001). Molecular Cloning: A Laboratory Manual, 3rd edn. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Sherlock, O., Dobrindt, U., Jensen, J. B., Munk Vejborg, R., and Klemm, P. (2006). Glycosylation of the self-recognizing Escherichia coli Ag43 autotransporter protein. J. Bacteriol. 188, 1798–1807. doi: 10.1128/JB.188.5.1798-1807.2006
Sherlock, O., Schembri, M. A., Reisner, A., and Klemm, P. (2004). Novel roles for the AIDA adhesin from diarrheagenic Escherichia coli: cell aggregation and biofilm formation. J. Bacteriol. 186, 8058–8065. doi: 10.1128/JB.186.23.8058-8065.2004
Sherlock, O., Vejborg, R. M., and Klemm, P. (2005). The TibA adhesin/invasin from enterotoxigenic Escherichia coli is self recognizing and induces bacterial aggregation and biofilm formation. Infect. Immun. 73, 1954–1963. doi: 10.1128/IAI.73.4.1954-1963.2005
Smith, D. R., Price, J. E., Burby, P. E., Blanco, L. P., Chamberlain, J., and Chapman, M. R. (2017). The production of curli amyloid fibers is deeply integrated into the biology of Escherichia coli. Biomolecules 7:E75. doi: 10.3390/biom7040075
Sugawara, E., and Nikaido, H. (1994). OmpA protein of Escherichia coli outer membrane occurs in open and closed channel forms. J. Biol. Chem. 269, 17981–17987.
Toguchi, A., Siano, M., Burkart, M., and Harshey, R. M. (2000). Genetics of swarming motility in Salmonella enterica serovar typhimurium: critical role for lipopolysaccharide. J. Bacteriol. 182, 6308–6321. doi: 10.1128/JB.182.22.6308-6321.2000
Tytgat, H. L., Van Teijlingen, N. H., Sullan, R. M., Douillard, F. P., Rasinkangas, P., Messing, M., et al. (2016). Probiotic gut microbiota isolate interacts with dendritic cells via glycosylated heterotrimeric pili. PLoS ONE 11:e0151824. doi: 10.1371/journal.pone.0151824
Valvano, M. A., Marolda, C. L., Bittner, M., Glaskin-Clay, M., Simon, T. L., and Klena, J. D. (2000). The rfaE gene from Escherichia coli encodes a bifunctional protein involved in biosynthesis of the lipopolysaccharide core precursor ADP-L-glycero-D-manno-heptose. J. Bacteriol. 182, 488–497. doi: 10.1128/JB.182.2.488-497.2000
Valvano, M. A., Messner, P., and Kosma, P. (2002). Novel pathways for biosynthesis of nucleotide-activated glycero-manno-heptose precursors of bacterial glycoproteins and cell surface polysaccharides. Microbiology 148, 1979–1989. doi: 10.1099/00221287-148-7-1979
Vandrangi, P., Lo, D. D., Kozaka, R., Ozaki, N., Carvajal, N., and Rodgers, V. G. (2013). Electrostatic properties of confluent Caco-2 cell layer correlates to their microvilli growth and determines underlying transcellular flow. Biotechnol. Bioeng. 110, 2742–2748. doi: 10.1002/bit.24939
Wessel, D., and Flügge, U. I. (1984). A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal. Biochem. 138, 141–143. doi: 10.1016/0003-2697(84)90782-6
Whitchurch, C. B., Tolker-Nielsen, T., Ragas, P. C., and Mattick, J. S. (2002). Extracellular DNA required for bacterial biofilm formation. Science 295, 1487. doi: 10.1126/science.295.5559.1487
Whitfield, C., and Trent, M. S. (2014). Biosynthesis and export of bacterial lipopolysaccharides. Annu. Rev. Biochem. 83, 99–128. doi: 10.1146/annurev-biochem-060713-035600
Keywords: enterotoxigenic Escherichia coli, Lipopolysaccharide, ADP-l-glycero-β-d-manno-heptose, HldE, biofilm formation, motility, transmission electron microscopy, virulence factors
Citation: Maigaard Hermansen GM, Boysen A, Krogh TJ, Nawrocki A, Jelsbak L and Møller-Jensen J (2018) HldE Is Important for Virulence Phenotypes in Enterotoxigenic Escherichia coli. Front. Cell. Infect. Microbiol. 8:253. doi: 10.3389/fcimb.2018.00253
Received: 13 April 2018; Accepted: 04 July 2018;
Published: 07 August 2018.
Edited by:
Philip R. Hardwidge, Kansas State University, United StatesReviewed by:
Jorge Blanco, Universidade de Santiago de Compostela, SpainMauricio J. Farfan, Universidad de Chile, Chile
George P. Munson, University of Miami, United States
Copyright © 2018 Maigaard Hermansen, Boysen, Krogh, Nawrocki, Jelsbak and Møller-Jensen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jakob Møller-Jensen, amFrb2JtQGJtYi5zZHUuZGs=