
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol., 28 June 2018
Sec. Molecular Bacterial Pathogenesis
Volume 8 - 2018 | https://doi.org/10.3389/fcimb.2018.00221
 Yousef Maali1
Yousef Maali1 Cédric Badiou1
Cédric Badiou1 Patrícia Martins-Simões1,2,3
Patrícia Martins-Simões1,2,3 Elisabeth Hodille1,2
Elisabeth Hodille1,2 Michele Bes2,3
Michele Bes2,3 François Vandenesch1,2,3
François Vandenesch1,2,3 Gérard Lina1,2,3
Gérard Lina1,2,3 Alan Diot1
Alan Diot1 Frederic Laurent1,2,3*†
Frederic Laurent1,2,3*† Sophie Trouillet-Assant1,2,4†
Sophie Trouillet-Assant1,2,4†Staphylococcus pseudintermedius is responsible for severe and necrotizing infections in humans and dogs. Contrary to S. aureus, the pathophysiological mechanisms involved in this virulence are incompletely understood. We previously showed the intracellular cytotoxicity induced after internalization of S. pseudintermedius. Herein, we aimed to identify the virulence factors responsible for this cytotoxic activity. After addition of filtered S. pseudintermedius supernatants in culture cell media, MG63 cells, used as representative of non-professional phagocytic cells (NPPc), released a high level of LDH, indicating that the cytotoxicity was mainly mediated by secreted factors. Accordingly, we focused our attention on S. pseudintermedius toxins. In silico analysis found the presence of two PSMs (δ-toxin and PSMε) as well as Luk-I leukotoxin, the presence of which was confirmed by PCR in all clinical strains tested (n = 17). Recombinant Luk-I leukotoxin had no cytotoxic activity on NPPc but the ectopic expression of the CXCR2 receptor in U937 cells conferred cytotoxity to Luk-I. This is in agreement with the lack of Luk-I effect on NPPc and the previous report of Luk-I cytoxic activity on immune cells. Contrary to Luk-I, synthetic δ-toxin and PSMε had a strong cytotoxic activity on NPPc. The secretion of δ-toxin and PSMε at cytotoxic concentrations by S. pseudintermedius in culture supernatant was confirmed by HPLC-MS. In addition, the supplementation of such supernatants with human serum, known to inhibit PSM, induced a complete abolition of cytotoxicity which indicates that PSMs are the key players in the cytotoxic phenotype of NPPc. The results suggest that the severity of S. pseudintermedius infections is, at least in part, explained by a combined action of Luk-I that specifically targets immune cells expressing the CXCR2 receptor, and PSMs that disrupt cell membranes whatever the cell types. The present study strengthens the key role of PSMs in virulence of the different species belonging to Staphylococcus genus.
Staphylococcus pseudintermedius is a skin and mucous commensal bacterium in dogs, with carriage reaching more than 80% in some populations of healthy dogs (Rubin and Chirino-Trejo, 2011; Bannoehr and Guardabassi, 2012). This species is also the most frequent bacterial pathogen in clinical canine specimens mainly from skin wounds but also the ears, bones or post-surgical abscesses (Fazakerley et al., 2009; Miedzobrodzki et al., 2010; Bannoehr and Guardabassi, 2012). In addition to this virulence profile, methicillin-resistance in S. pseudintermedius (MRSP) has emerged worldwide over the two last decades and the treatment of S. pseudintermedius infections is now considered as a clinical challenge in veterinary medicine. In addition, S. pseudintermedius is increasingly isolated in human infections, mainly after contacts with dogs (Somayaji et al., 2016). Thus, since the first human case was described in 2006 (endocarditis occurring after the implantation of a cardioverter-defribrillator device Van Hoovels et al., 2006), sporadic community-acquired infections as well as nosocomial outbreaks involving S. pseudintermedius have been reported (Starlander et al., 2014; Somayaji et al., 2016).
Whether from canine or human origin, S. pseudintermedius infections are classically severe due to necrotizing processes, as illustrated in dermatitis and fasciitis in which a deep destruction of cells and tissues are observed (Weese et al., 2009). S. pseudintermedius, often described as the “dog's golden staph,” shares several features with S. aureus, notably the capacity to express a variety of virulence factors such as: (i) proteolytic enzymes including coagulase and protease (Garbacz et al., 2013), (ii) microbial surface components recognizing adhesive matrix molecules (MSCRAMMs), such as the staphylococcal protein A, that are host surface proteins and participate to the active tissue colonization and the evasion of the host immune system (Bannoehr et al., 2011, 2012; Balachandran et al., 2017), and (iii) toxins (Dziewanowska et al., 1996; Futagawa-Saito et al., 2004). Among the latter, pore-forming toxins (PFTs) have been extensively investigated for their ability to damage plasma membranes and lead to cell lysis (Prévost et al., 2001). Some of them induce cells death after the binding of the toxin with receptors on cell surface. This is the case for S. aureus bicomponent leukotoxins such as the Panton–Valentine leukocidin (PVL) or the leukotoxin ED (LukED) which act only on immune cells because of the interaction with cell receptors C5a or CXCR2, respectively (Reyes-Robles et al., 2013; Spaan et al., 2013). S. pseudintermedius produces a bicomponent leukotoxin, Luk-I, composed of the secreted LukS-I and LukF-I proteins that induce cell lysis. This activity has been reported only on PMNs and to date, no receptor has yet been identified (Prevost et al., 1995). Recently, in vitro studies underlined the highly virulent behavior of S. pseudintermedius on non-professional phagocytic cells (NPPc) such as canine keratinocytes, human epithelial cells and human osteoblasts, which are the most impacted cells during S. pseudintermedius infections (Pietrocola et al., 2015; Maali et al., 2016). Pietrocola et al. suggested that the S. pseudintermedius bicomponent leukotoxin Luk-I could also be involved in the cell death observed after internalization of S. pseudintermedius in NPPc (Pietrocola et al., 2015). To the best of our knowledge, this hypothesis has yet to be investigated. Phenol-soluble modulins (PSMs) represent another class of membrane damaging staphylococcal toxins. This class of amphipathic proteins has alpha-helical structure, and has a direct and receptor-independent action on the lipid bilayers of cells (Peschel and Otto, 2013). Various types of PSMs (α, β, δ-toxin…) have been described and are highly conserved among S. aureus or Staphylococcus non-aureus (SNA) strains (Cheung et al., 2014; Cameron et al., 2015; Da et al., 2017); whole genome sequencing analysis of S. pseudintermedius has revealed the presence of several PSMs: δ-toxin, PSMβ, and PSMε (Cheung et al., 2014).
Taking into consideration all of these elements, we sought to identify the virulence factors involved in the cytolytic effect on NPPc and immune cells related to severe and necrotizing S. pseudintermedius infections. Using in vitro cellular models (immune cells, NPPc, and transfected cells) and purified synthetic or recombinant toxins, bacterial culture supernatants with or without PSMs inhibitors; we identified the membrane receptor of Luk-I present on immune cells and demonstrated the specific S. pseudintermedius cytolytic activity of PSMε and δ-toxin on both NPPc and immune cells.
This study was approved by the French South-East ethics committee (reference number 2013-018). In accordance with French legislation, written informed patient consent was not required for the use of the collected clinical isolates.
A collection of 17 human and animal clinical S. pseudintermedius isolates was used. Characteristics of the isolates are summarized in Table 1. Identifications of all strains were confirmed using matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF-MS; VITEK® MS, bioMérieux, Marcy-L'Etoile, France) (Silva et al., 2015). The strains S. aureus LAC USA300 and its isogenic mutant S. aureus LAC Δpsm (α, β, δ), kindly provided by Frank Deleo (National Institutes of Health, Hamilton, MT, US) were used as positive and negative controls, respectively, for cytotoxicity experiments (Wang et al., 2007; Davido et al., 2016).
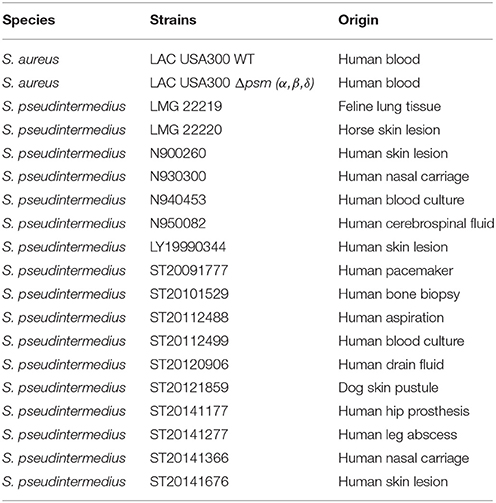
Table 1. Description of the strains used.
Screening for the presence of lukF-I and lukS-I genes encoding leukocidin Luk-I, psmε that encodes PSMε, and hld that encodes δ-toxin was performed in the S. pseudintermedius isolates by PCR. Oligonucleotides and annealing temperatures are presented in the Supplementary Table 1. The resulting PCR products were sequenced (Biofidal, Lyon, France) and aligned using SeaView (Version 4.6.3; PRABI, Lyon, France) to investigate the genetic polymorphism of each gene (Gouy et al., 2010).
Purified toxins were then obtained by recombinant or synthetic production. For Luk-I, lukS-I and lukF-I genes were amplified by PCR using chromosomal DNA of S. pseudintermedius LMG 22219 using flanking primers, the sequences of which are presented in Supplementary Table 1. PCR products were codigested with BamHI and SalI enzymes (New England Biolabs, Ipswich, MA,US) then purified using the High Pure PCR Product Purification kit (Roche, Meylan, France) and ligated using T4 DNA Ligase (Promega, Madison, WI, USA) into the pQE-30 plasmid (Qiagen, Courtaboeuf, France). The resulting pQE-lukS-I and pQE-lukF-I plasmids were transformed into E. coli strain M15. Recombinant His-tagged toxins were expressed in LB medium and purified with Ni-nitrilotriacetic acid columns (Qiagen) before being dialyzed against phosphate-buffered saline (PBS). After protein purity was checked using SDS-PAGE and protein quantification performed using the Bradford method, LukS-I and LukF-I were combined and used for the cytotoxicity assays described below.
For PSMs, amino acid sequences corresponding to PSMε and the two variants of δ-toxin (variant ED99 and variant HKU10-03) were synthetized and purified to >90% by Genecust (Dudelange, Luxembourg) using high-performance liquid chromatography (HPLC).
Staphylococcal supernatants of tested strains were prepared by growing staphylococci bacteria in brain heart infusion (BHI) medium (bioMérieux) in a rotary shaker (190 rpm) at 37°C for 22 h, followed by centrifugation for 10 min at 3,000 × g. The filtered supernatants were stored at −80°C before being tested. For PSM quantification, bacterial supernatants were diluted 1:5 in methanol at 4°C, and incubated for 10 min at 4°C for cold precipitation to eliminate particulate matter. After centrifugation at 10,000 × g for 5 min, PSMε and δ-toxin (variant ED99 and variant HKU10-03) were quantified using a targeted approach by HPLC—mass spectrometry (HPLC-MS) in an Agilent® system as described previously (Hodille et al., 2016).
All cell culture reagents were obtained from Gibco (Paisley, United Kingdom). Human MG63 osteoblastic cells (LGC Standards, Teddington, UK), an NPPc cell line, were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 2 mM L-glutamine and 25 mM HEPES and supplemented with 10% heat-inactivated fetal bovine serum (FBS) as well as 100 U/mL penicillin and streptomycin. Cells were passaged twice a week and were used in experiments up to passage 20 after being thawed from a stock culture.
U937 cells (human promyelocytic cell line) and U937 cells stably transfected with human C5a receptor (U937-C5aR), or with the CXCR2 (U937-CXCR2) kindly provided by Jos A. G. Van Strijp (Department of Medical Microbiology, Utrecht, Netherlands), were cultured at 37°C, 5% CO2 in Roswell Park Memorial Institute medium (RPMI) 1640 containing 10% heat-inactivated FBS (Kew et al., 1997).
Human polymorphonuclear neutrophils (PMNs) collected from the blood of healthy donors were isolated by density gradient centrifugation on Ficoll/Histopaque (Sigma-Aldrich) (Veldkamp et al., 2000). Viability of PMNs was determined by trypan blue exclusion and ranged from 95 to 99%. PMN were maintained at 37°C, 5% CO2 in RPMI 1640 containing 10% heat-inactivated FBS.
The cytotoxicity induced by the staphylococcal toxins was evaluated by measuring propidium iodide (PI) incorporation in to cells exposed to toxins using a fluorescence cell sorter (Spark®, TECAN, Zürich, Switzerland). Briefly, MG63 osteoblasts were seeded at 104 cells per well on 96-well tissue culture plates (Falcon, Le Pont de Claix, France) in 100 μL of culture media. After 24 h, the cells were washed three times with 100 μL of PBS (Gibco). Immune cells (U937 and PMN) at 105 cells per well were seeded in tissue culture plates the day of the experiment. All cells were incubated for 3 h at 37°C with toxins and PI (1.25 μg.mL−1). For the Luk-I toxin, 0.5 to 5 μg.mL−1 were tested after dilution in culture medium. For PSMε and the two δ-toxin variants, 0.1 to 100 μg.mL−1 were tested after dilution in DMEM (Flammier et al., 2016).
The negative control (Cneg) corresponded to the same experiment without toxin. The positive control (Cpos) was obtained using complete cell lysis solution (Triton X100 0.1%). As previously described, cell lysis due to toxins was expressed as percentages using the following formula: (Hodille et al., 2016).
Because of the high autofluorescence of BHI medium, the cytotoxic effect of culture supernatant on NPPc could not be performed using IP incorporation. To circumvent this issue, cytotoxicity was evaluated by measuring Lactate dehydrogenase (LDH) release in the cell culture supernatant using a colorimetric method (Dimension Vista automated clinical chemistry analyzer, Siemens Healthcare Diagnostics, Tarrytown, NY, USA). In parallel, the cytotoxic effect of the supernatant was investigated by microscopy; after being fixed using formaldehyde 4% for 20 min, and stained 30 min using Giemsa stain, modified solution (Sigma-Aldrich) according to the manufacturer's instructions, cells were observed under a light microscope Leica DMi1 (Leica Microsystems, Wetzlar, Germany) at a magnification of × 20. The supernatant from cytotoxic S. aureus LAC USA300 strain was used as positive control, and sterile BHI medium was used as negative control.
Osteoblasts were seeded at 100,000 cells per well on 24-well tissue culture plates (Falcon) in 1 mL of culture media. After 24 h, the cells were washed twice with 1 mL of DMEM before the addition of filter-sterilized (0.22 μm) staphylococcal culture supernatants. Serum inhibition of PSM cytotoxic activity in supernatant was tested using pooled human serum obtained from healthy donors provided by the French national public blood service (Etablissement Français du Sang, Lyon, France). Filter-sterilized supernatants were mixed with or without 5% inactivated human serum for 10 min at room temperature (Surewaard et al., 2012). Then, NPPC (osteoblasts) were incubated for 4 h at 37°C, 5% CO2.
Non-parametric statistical Mann-Whitney U-tests and B tests with a threshold of significance of 0.05 were used to determine statistically significant differences. All analyses were performed using XLSTAT software (Pearson Edition, developed by Addinsoft v.2014; New York, NY, US).
Knowing that the supernatant of S. pseudintermedius is able to induce the death of human PMNs (Riegel et al., 2011), we specifically performed an in vitro cytotoxic assay to evaluate its capacity to also kill non-immune cells, such as NPPc. First, cytolytic activity expressed as the means ± standard deviations quantified by LDH release from osteoblasts incubated with S. pseudintermedius LMG 20220 culture supernatant (773 ± 92%), was significantly higher than those observed in the control condition (100 ± 12%, p < 0.001; Figure 1A). Like S. pseudintermedius, S. aureus culture supernatant has a high cytotoxic effect on NPPc. Second, the microscopic observations indicated a high cytotoxic action of the supernatant from S. pseudintermedius LMG 20220 on MG63 cells, as demonstrated by the lysis area compared to the uniform cell layer observed in the control condition; the same phenotypic profile was observed for the hyper virulent strain S. aureus LAC USA300 (Figure 1B). These results demonstrate that the cytotoxicity induced by S. pseudintermedius is mediated by secreted virulence factors, which prompted us to investigate the specific bacterial factors involved, especially among staphylococcal toxins with cellular tropism.
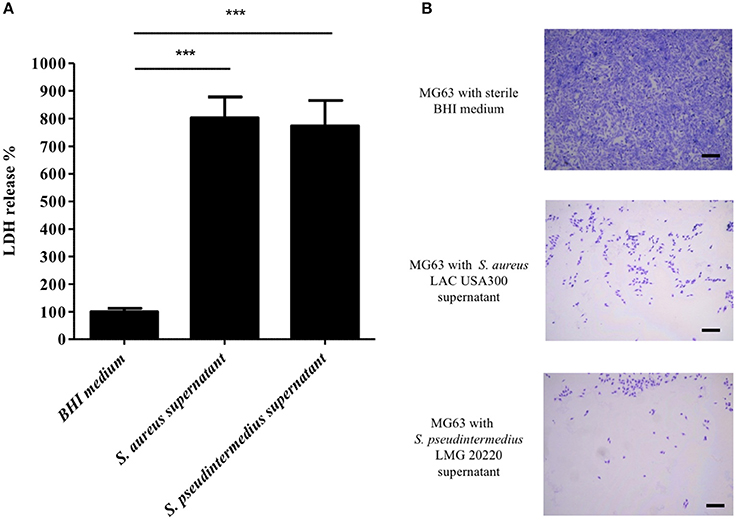
Figure 1. Effect of S. pseudintermedius supernantant on NPPc (MG63). MG63 cells were incubated with staphylococcal supernatants for 4 h at 37°C. (A) Quantification of LDH release, reflecting NPPc lysis by staphylococcal supernatant. All of the results are expressed as the percentages of the values obtained for the control (BHI medium alone, 100%). Bars represent means ± standard deviations derived from 3 experiments performed in triplicate. The difference in the LDH concentration in staphylococcal supernatants condition compared to BHI medium alone was evaluated using a one-tailed Mann-Whitney test with a α risk of 0.05 (***p < 0.001). (B) MG63 lysis after 4 h in contact with staphylococcal supernatant. The cells were stained with Giemsa, and observed for morphological changes by light microscopy at a magnification of ×20. Bars, 500 μm. LDH, Lactate dehydrogenase; NPPc, Non-professional phagocytic cells.
PCR experiments found that all S. pseudintermedius isolates were positive for lukF-I / lukS-I genes. The use of recombinant Luk-I (combining LukS-I and LukF-I) allowed us to specifically analyze in vitro the cytotoxic capacity of this toxin. PMNs were used as control because the activity of Luk-I on these cells has been previously described (Prevost et al., 1995). As expected, a very high cytotoxic activity was observed for the PMNs cells, and this was found at the lowest concentration of recombinant Luk-I tested (0.5 μg.mL−1; Figure 2A). Conversely, the same toxin had no cytotoxic activity against NPPc for all tested concentrations (from 0.5 to 5 μg.mL−1). In order to identify the mechanism of this specific activity for PMNs, we tested the activity of toxins on wild-type U937 cells and transfected U937 cells that express the C5a or CXCR2 receptors; Luk-I had a high cytotoxic action only on cells that expressed the CXCR2 receptor (Figure 2B). This result suggests that the toxin Luk-I exhibits a tropism for myeloid cells expressing the CXCR2 receptor.
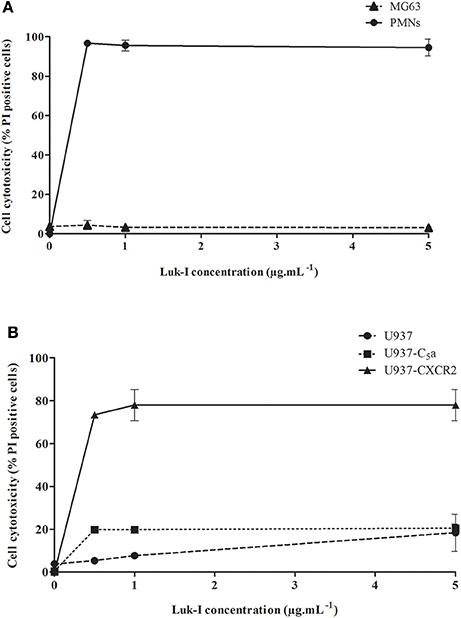
Figure 2. Cytolytic activity of S. pseudintermedius leukotoxin Luk-I. (A) Leukotoxin Luk-I toward NPPc (MG63) and immune cells (human PMNs). (B) Leukotoxin Luk-I activity on wild-type U937 cells and U937 cells expressing the C5a or CXCR2 receptors. Cells were incubated with increasing doses of recombinant Luk-I (from 0.5 to 5 μg.mL−1) for 3 h at 37°C. Cell death was measured by staining cells with propidium iodide (PI) and detected with a fluorescence cell sorter. The percentage of cell lysis was calculated as . The values represent the means ± standard deviations derived from one experiment performed in triplicate representative of two others. NPPc, Non-professional phagocytic cells; PMNs, Polymorphonuclear neutrophils; PI, Propidium iodide; U937-C5a, U937 cells transfected with C5a receptor; U937-CXCR2, U937 cells transfected with CXCR2 receptor.
Due to the absence of activity for Luk-I on NPPc (MG63 cells), we speculated that PSMs could be the key virulence factors implicated in the S. pseudintermedius virulence observed on these cells. Two PSMs with cellular tropisms have been identified in S. pseudintermedius using in silico analysis based on the NCBI database: PSMε and the δ-toxin, for which two variants (ED99 and HKU10-03) have been described (Cheung et al., 2014). We therefore screened for the presence of these PSMs in the collection of clinical strains. Sequencing of the PCR products found the presence of psmε and hld in all S. pseudintermedius isolates (n = 17). For δ-toxin, 14/17 isolates (82%) harbored the variant ED99 while 3/17 (18%) harbored the variant HKU10-03 (Table 2).

Table 2. Amino acid sequence of S. pseudintermedius phenol-soluble modulins (PSM).
To investigate the expression of these 3 PSMs, we quantified the level of their secretion in the supernatant of strains S. pseudintermedius LMG 20220 and S. pseudintermedius ST20112488, which respectively secrete δ-toxin variant ED99 and variant HKU10-03 according to the genetic profile (Supplementary Figure 1). Both PSMε and the two δ-toxin variants were detected and distinguished by HPLC-MS, and the mean ± SD concentrations in culture supernatants ranged from 22.4 ± 2.8 μg.mL−1 to 159.3 ± 83.9 μg.mL−1 (Table 3).
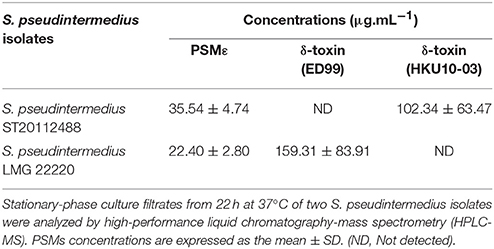
Table 3. Phenol-soluble modulins (PSM) concentrations in S. pseudintermedius supernatants.
In order to evaluate the cytotoxic effect of PSMs that we identified in the collection of S. pseudintermedius isolates, synthetic PSMε and δ-toxin variants covering the ranges of secreted concentrations were tested in an in vitro model (Figure 3). All types of PSMs tested were cytotoxic for NPPc in a dose-dependent manner. Knowing that PSMs have a receptor-independent and direct action on the lipid bilayers of cells, as expected, human PMNs were also sensitive to the different S. pseudintermedius PSMs (Supplementary Figure 2).
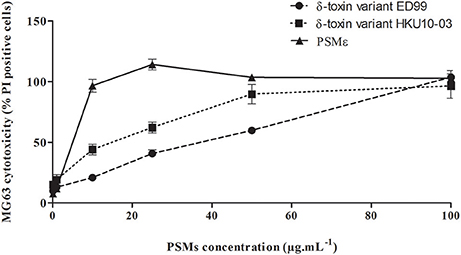
Figure 3. Cytolytic activity of S. pseudintermedius PSMs. MG63 cells were incubated with increasing concentrations (from 0.1 to 100 μg.mL−1) of synthetic PSMε, δ-toxin (ED99) or δ-toxin (HKU10-03) for 3 h at 37°C. Cell death was measured by staining cells with propidium iodide and detected with a fluorescence cell sorter. The percentage of cell lysis was calculated as . The values represent the means ± standard deviations derived from one experiment performed in triplicate representative of two others. NPPc, Non-professional phagocytic cells; PMNs, Polymorphonuclear neutrophils; PSM, Phenol-soluble modulins; PI, Propidium iodide.
To confirm the hypothesis that the virulence observed on the NPPc was mediated mainly by the PSMs secreted by S. pseudintermedius, a serum inhibition assay was conducted firstly on S. aureus LAC USA300 strains and its isogenic mutant Δpsm (α, β, δ) (Figure 4). LDH release results were expressed as percentages of control condition (100%, medium alone). A complete abolition of cytotoxicity capacity of S. aureus LAC USA300 supernatant was found after the addition of human serum (LDH release - 99.02 ± 7.14%). No significant difference was observed between S. aureus Δpsm (α, β, δ) supernatant in the presence or not of human serum. This result confirmed the specific effect of human serum on PSM activity. Secondly, similarly to that found with S. aureus LAC USA300, the cytolytic activity of the strains S. pseudintermedius LMG 22220 (expressing PSMε and δ-toxin variant ED99) and S. pseudintermedius ST20112488 (expressing PSMε and δ-toxin variant HKU10-03) were completely abolished in presence of human serum addition (respectively, 1097.22 ± 143.84% vs. 106.71 ± 14.84%, p < 0.001 and 1081.94 ± 224.04% vs. 96.68 ± 11.71%, p < 0.001). Moreover, the cytotoxicity induced by synthetic PSMs peptides was completely inhibited in presence of inactivated human serum whereas Luk-I activity is not affected (Supplementary Figure 3). The data indicates the preponderant PSM action in the cytotoxicity of NPPc and consequently the key role of these in the virulence of S. pseudintermedius.
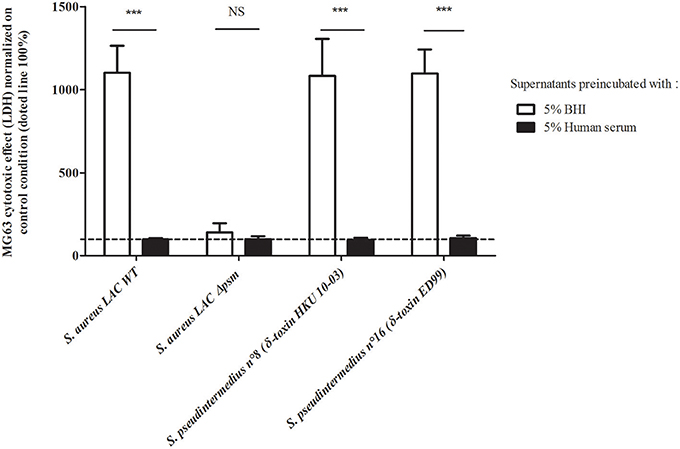
Figure 4. Human serum action on staphylococcal culture supernatants. MG63 cells were incubated for 4 h at 37°C with staphylococcal supernatants preincubated with 5% heat-inactivated human serum or with 5% BHI. Results are expressed as the percentages of the values obtained for the control condition “medium supplemented with BHI or human serum 5%” (100%), represented by the dotted line. Bars represent means ± standard deviations derived from three experiments performed in triplicate. The difference in the LDH concentration in staphylococcal supernatants condition compared to the uninfected control cells was evaluated using a one-tailed Mann-Whitney test with an α risk of 0.05 ***p < 0.001; NS, Non-significant; BHI, Brain heart infusion; LDH, Lactate dehydrogenase; NPPc, Non-professional phagocytic cells.
S. pseudintermedius is an opportunistic pathogen responsible for particularly severe infection both in humans and dogs. The severity of these infections has been illustrated by several clinical cases including cellulitis, purulent exudate, inflammation, and necrosis (Weese et al., 2009; Stegmann et al., 2010; Riegel et al., 2011; Mayer and Rubin, 2012; Lee et al., 2015; Darlow et al., 2017; Robb et al., 2017). Contrary to S. aureus, for which a panel of virulence factors have been identified and characterized in the various pathophysiological contexts, knowledge about the pathogenesis of S. pseudintermedius remains limited. Through in vitro assays, the present study allowed us to identify the key players of the high cellular virulence associated with S. pseudintermedius infections.
The study found that the supernatant of S. pseudintermedius had a strong cytotoxic effect on NPPc suggesting that this cytotoxicity is mediated, at least in part, by secreted virulence factors. Several authors have suggested that Luk-I (present in all S. pseudintermedius isolates tested) could be responsible for cell death induction in NPPc (Garbacz et al., 2013; Pietrocola et al., 2015). Herein, no cytotoxic activity was observed with the recombinant Luk-I toxin against NPPc over the range of concentrations tested, conversely to that found for PMN cells. The lack of Luk-I activity on NPPc led us to draw parallels with S. aureus leukotoxins that induce cell lysis only on immune cells harboring specific receptors. To address this question, we performed an in vitro assay with U937 promyelocytic cells stably transfected with C5a and CXCR2 receptors, which are respectively the target receptors of PVL and LukED leukotoxin. The results indicate that Luk-I induces cell death via the CXCR2 receptor specifically present on immune cells (PMNs, macrophages) as previously reported for leukotoxin LukED for S. aureus, explaining the absence of cytotoxicity observed in NPPc. This feature is likely related to the high homology between Luk-I and LukED (Spaan et al., 2017). Interestingly, this receptor, found both in humans and dogs, also shares a high homology (75%) (Hall et al., 2010).
Wang et al. have reported the contribution of a family of pro-inflammatory and cytolytic staphylococcal peptide toxins, called PSMs, in several disease manifestations (Wang et al., 2007). Moreover, Cheung et al. reported that PSMs δ, β, ε were present in the genome of S. pseudintermedius through in silico studies (Cheung et al., 2014). Herein, psmε and hld genes, encoding for the two PSMs described in S. pseudintermedius able to induce cytolysis, were present in all clinical isolates (Table 1) (Otto, 2014). Analysis by HPLC-MS of δ-toxin and PSMε showed that these toxins were secreted at a high level (up to 100 μg.mL−1) by clinical S. pseudintermedius strains in in vitro conditions. Regarding the level of PSMs secreted in vivo, to the best of our knowledge, only one study has reported data on S. aureus; on in vivo samples of human pus, in which PSMs concentrations were detected up to 20 μg.mL−1 (Hodille et al., 2016). Using synthetic peptides, we found that all S. pseudintermedius PSMs studied herein are able to induce osteoblast death in a dose-dependent manner. These results confirm the cytolytic action of α-class PSMs for which it is well established in the literature that these peptides can disrupt plasma membranes independently of cell type (Wang et al., 2007; Otto, 2014). Because most α-type PSMs are sometimes too short to be annotated in genome sequences and to yield significant results in similarity analysis, in silico studies do not allow the detection of all α-type psm genes in a given genome. It would be of interest to more deeply explore the potential presence of other PSMs involved in the virulence of S. pseudintermedius.
The construction of isogenic mutant Δpsm would have been the classical way to definitively confirm that PSMs are the main factors responsible for S. pseudintermedius cytotoxicity on NPPc. Unfortunately, isogenic gene deletion mutants are extremely difficult to obtain in this species, which is further compounded by the multiplicity of PSMs and their functional redundancy, making it necessary to produce sequential, multiple knockouts for psm gene loci; such a construction was unsuccessful (data not shown). Faced with this technical limit, we chose another strategy which involved the addition of human serum that is known to be specific inhibitors of PSM action (Surewaard et al., 2012). This led to a complete loss of the cytotoxicity effect of S. pseudintermedius supernatant, attesting to the role of PSMs in the virulence of S. pseudintermedius.
The results of the present study reinforce the central role of PSMs in the virulence of staphylococci. These molecules are widespread among staphylococcal species and participate in pleiotropic action such as cytolysis (Rasigade et al., 2013; Da et al., 2017), surface colonization (Tsompanidou et al., 2013), biofilm structure and biofilm detachment (Wang et al., 2011). S. pseudintermedius is preferentially involved in skin and soft tissue infections with muco-purulent exudates, environments in which these amphipathic toxins retain all their active properties. In addition, S. pseudintermedius presents the rare capacity to internalize into host cells, therefore the intracellular compartment also constitutes a microenvironment for the accumulation of PSMs and protection from serum lipoproteins (Rasigade et al., 2013). Virulence of S. pseudintermedius is explained mainly by PSMs acting both in the extracellular and intracellular compartments of cells and by a selective action of Luk-I toxins acting on immune cells expressing CXCR2 receptors (Figure 5).
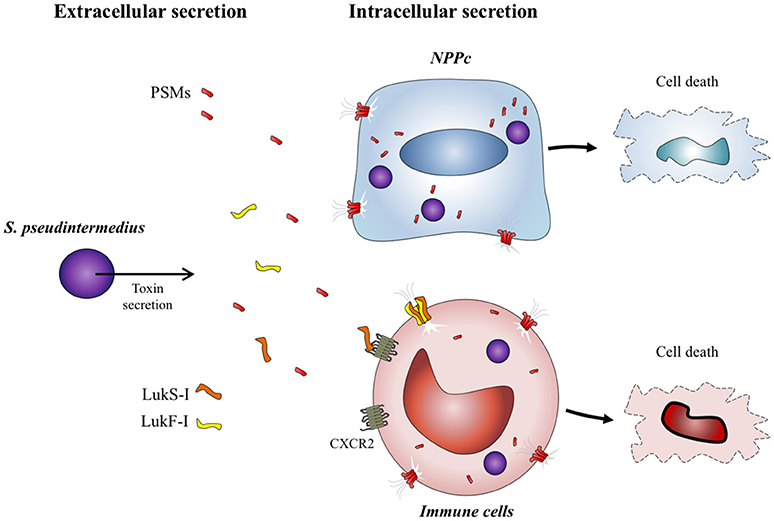
Figure 5. S. pseudintermedius toxins action on host cells. S. pseudintermedius can cause the necrosis of immune cells such as human polymorphonuclear neutrophils (PMNs) through the release of bicomponent leukotoxin Luk-I which recognizes the CXCR2 receptor. S. pseudintermedius is also able to secrete other membrane-damaging virulence factors: the phenol-soluble modulins (PSMs) which have a cytotoxic effect on both immune cells and NPPc.
The present study helps elucidate bacterial pathophysiological mechanisms that could explain the severity of human and animal infections related to S. pseudintermedius which behaves similarly to S. aureus. Both harbor a variety of common virulence factors including similar/homologous MSCRAMMs, bi-component leukotoxin and PSMs, and both species are able to induce clinically severe infections. In vitro, they exhibit the same behavior toward human cells: they are similarly able to attack, invade and lyse human cells. Interestingly, while S. pseudintermedius infections are highly prevalent in pets, especially in dogs, in which it colonizes in particular mucocutaneous sites, such as the nose, mouth and anus with a carriage rate that reaches 92% in some studies, human S. pseudintermedius infections are scarce and classically occur after contact with animals and in presence of skin barrier defects (Allaker et al., 1992a,b; Rubin and Chirino-Trejo, 2011; Bannoehr and Guardabassi, 2012). This could be explained by the carriage rate in humans, which has been demonstrated to be very low (Paul et al., 2011; Trouillet-Assant et al., 2015). In contrast, S. aureus infections are very common in humans and related to preexisting nasal carriage (self-contamination) or to transmission from colonized health caregivers. This suggests that the host specificity of the two species might be related to the specific colonizing capacity of each species. Further studies deserve to be conducted to explore this differential ability and to identify the mechanisms.
FL and ST-A: conceived and designed the experiments; YM and CB: performed the experiments; YM: data collection; YM, CB, PM-S, and EH: data analysis; MB, FV, AD, GL, ST-A, and FL: data interpretation; YM and ST-A: drafting manuscript; CB, EH, MB, GL, AD, ST-A, and FL: revising manuscript content; FL: takes responsibility for the integrity of the data analysis.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was supported by the Institut National de la Santé et de la Recherche Médicale (INSERM) and Allocations de Recherche du Ministère de la Recherche. We thank Dr. Yvonne Benito for her generous technical assistance on molecular biology. We thank Dr. Karen Moreau for her help on the Light Microscopy (Leica DMi1). We thank Dr. Carla de Haas in Utrecht for the cell lines. We thank Philip Robinson for language editing and critical reading the manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2018.00221/full#supplementary-material
Allaker, R. P., Lloyd, D. H., and Bailey, R. M. (1992a). Population sizes and frequency of staphylococci at mucocutaneous sites on healthy dogs. Vet. Rec. 130, 303–304. doi: 10.1136/vr.130.14.303
Allaker, R. P., Lloyd, D. H., and Simpson, A. I. (1992b). Occurrence of Staphylococcus intermedius on the hair and skin of normal dogs. Res. Vet. Sci. 52, 174–176. doi: 10.1016/0034-5288(92)90006-N
Balachandran, M., Bemis, D. A., and Kania, S. A. (2017). Expression and function of protein A in Staphylococcus pseudintermedius. Virulence 9, 390–401. doi: 10.1080/21505594.2017.1403710
Bannoehr, J., Ben Zakour, N. L., Reglinski, M., Inglis, N. F., Prabhakaran, S., Fossum, E., et al. (2011). Genomic and surface proteomic analysis of the canine pathogen Staphylococcus pseudintermedius reveals proteins that mediate adherence to the extracellular matrix. Infect. Immun. 79, 3074–3086. doi: 10.1128/IAI.00137-11
Bannoehr, J., Brown, J. K., Shaw, D. J., Fitzgerald, R. J., van den Broek, A. H., and Thoday, K. L. (2012). Staphylococccus pseudintermedius surface proteins SpsD and SpsO mediate adherence to ex vivo canine corneocytes. Vet. Dermatol. 23, 119–24, e26. doi: 10.1111/j.1365-3164.2011.01021.x
Bannoehr, J., and Guardabassi, L. (2012). Staphylococcus pseudintermedius in the dog: taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet. Dermatol. 23, 253–66, e51-2. doi: 10.1111/j.1365-3164.2012.01046.x
Cameron, D. R., Jiang, J. H., Hassan, K. A., Elbourne, L. D., Tuck, K. L., Paulsen, I. T., et al. (2015). Insights on virulence from the complete genome of Staphylococcus capitis. Front. Microbiol. 6:980. doi: 10.3389/fmicb.2015.00980
Cheung, G. Y., Joo, H. S., Chatterjee, S. S., and Otto, M. (2014). Phenol-soluble modulins–critical determinants of staphylococcal virulence. FEMS Microbiol. Rev. 38, 698–719. doi: 10.1111/1574-6976.12057
Da, F., Joo, H. S., Cheung, G. Y. C., Villaruz, A. E., Rohde, H., Luo, X., et al. (2017). Phenol-soluble modulin toxins of Staphylococcus haemolyticus. Front. Cell. Infect. Microbiol. 7:206. doi: 10.3389/fcimb.2017.00206
Darlow, C. A., Paidakakos, N., Sikander, M., and Atkins, B. (2017). A spinal infection with Staphylococcus pseudintermedius. BMJ Case Rep. 2017:bcr-2017-221260. doi: 10.1136/bcr-2017-221260
Davido, B., Saleh-Mghir, A., Laurent, F., Danel, C., Couzon, F., Gatin, L., et al. (2016). Phenol-soluble modulins contribute to early sepsis dissemination not late local USA300-osteomyelitis severity in rabbits. PLoS ONE 11:e0157133. doi: 10.1371/journal.pone.0157133
Dziewanowska, K., Edwards, V. M., Deringer, J. R., Bohach, G. A., and Guerra, D. J. (1996). Comparison of the beta-toxins from Staphylococcus aureus and Staphylococcus intermedius. Arch. Biochem. Biophys. 335, 102–108. doi: 10.1006/abbi.1996.0486
Fazakerley, J., Nuttall, T., Sales, D., Schmidt, V., Carter, S. D., Hart, C. A., et al. (2009). Staphylococcal colonization of mucosal and lesional skin sites in atopic and healthy dogs. Vet. Dermatol. 20, 179–184. doi: 10.1111/j.1365-3164.2009.00745.x
Flammier, S., Rasigade, J. P., Badiou, C., Henry, T., Vandenesch, F., Laurent, F., et al. (2016). Human monocyte-derived osteoclasts are targeted by staphylococcal pore-forming toxins and superantigens. PLoS ONE 11:e0150693. doi: 10.1371/journal.pone.0150693
Futagawa-Saito, K., Sugiyama, T., Karube, S., Sakurai, N., Ba-Thein, W., and Fukuyasu, T. (2004). Prevalence and characterization of leukotoxin-producing Staphylococcus intermedius in Isolates from dogs and pigeons. J. Clin. Microbiol. 42, 5324–5326. doi: 10.1128/JCM.42.11.5324-5326.2004
Garbacz, K., Zarnowska, S., Piechowicz, L., and Haras, K. (2013). Pathogenicity potential of Staphylococcus pseudintermedius strains isolated from canine carriers and from dogs with infection signs. Virulence 4, 255–259. doi: 10.4161/viru.23526
Gouy, M., Guindon, S., and Gascuel, O. (2010). SeaView version 4: a multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 27, 221–224. doi: 10.1093/molbev/msp259
Hall, J. A., Chinn, R. M., Vorachek, W. R., Gorman, M. E., and Jewell, D. E. (2010). Aged Beagle dogs have decreased neutrophil phagocytosis and neutrophil-related gene expression compared to younger dogs. Vet. Immunol. Immunopathol. 137, 130–135. doi: 10.1016/j.vetimm.2010.05.002
Hodille, E., Cuerq, C., Badiou, C., Bienvenu, F., Steghens, J. P., Cartier, R., et al. (2016). Delta hemolysin and phenol-soluble modulins, but not alpha hemolysin or panton-valentine leukocidin, induce mast cell activation. Front. Cell. Infect. Microbiol. 6:180. doi: 10.3389/fcimb.2016.00180
Kew, R. R., Peng, T., DiMartino, S. J., Madhavan, D., Weinman, S. J., Cheng, D., et al. (1997). Undifferentiated U937 cells transfected with chemoattractant receptors: a model system to investigate chemotactic mechanisms and receptor structure/function relationships. J. Leukoc. Biol. 61, 329–337. doi: 10.1002/jlb.61.3.329
Lee, J., Murray, A., Bendall, R., Gaze, W., Zhang, L., and Vos, M. (2015). Improved detection of Staphylococcus intermedius group in a routine diagnostic laboratory. J. Clin. Microbiol. 53, 961–963. doi: 10.1128/JCM.02474-14
Maali, Y., Martins-Simoes, P., Valour, F., Bouvard, D., Rasigade, J. P., Bes, M., et al. (2016). Pathophysiological mechanisms of staphylococcus non-aureus bone and joint infection: interspecies homogeneity and specific behavior of S. pseudintermedius. Front. Microbiol. 7:1063. doi: 10.3389/fmicb.2016.01063
Mayer, M. N., and Rubin, J. E. (2012). Necrotizing fasciitis caused by methicillin-resistant Staphylococcus pseudintermedius at a previously irradiated site in a dog. Can. Vet. J. 53, 1207–1210.
Miedzobrodzki, J., Kasprowicz, A., Bialecka, A., Jaworska, O., Polakowska, K., Wladyka, B., et al. (2010). The first case of a Staphylococcus pseudintermedius infection after joint prosthesis implantation in a dog. Pol. J. Microbiol. 59, 133–135.
Otto, M. (2014). Phenol-soluble modulins. Int. J. Med. Microbiol. 304, 164–169. doi: 10.1016/j.ijmm.2013.11.019
Paul, N. C., Moodley, A., Ghibaudo, G., and Guardabassi, L. (2011). Carriage of methicillin-resistant Staphylococcus pseudintermedius in small animal veterinarians: indirect evidence of zoonotic transmission. Zoon. Public Health 58, 533–539. doi: 10.1111/j.1863-2378.2011.01398.x
Peschel, A., and Otto, M. (2013). Phenol-soluble modulins and staphylococcal infection. Nat. Rev. Microbiol. 11, 667–673. doi: 10.1038/nrmicro3110
Pietrocola, G., Gianotti, V., Richards, A., Nobile, G., Geoghegan, J. A., Rindi, S., et al. (2015). Fibronectin binding proteins spsd and spsl both support invasion of canine epithelial cells by Staphylococcus pseudintermedius. Infect. Immun. 83, 4093–4102. doi: 10.1128/IAI.00542-15
Prevost, G., Bouakham, T., Piemont, Y., and Monteil, H. (1995). Characterisation of a synergohymenotropic toxin produced by Staphylococcus intermedius. FEBS Lett. 376, 135–140. doi: 10.1016/0014-5793(95)01260-9
Prévost, G., Mourey, L., Colin, D. A., and Menestrina, G. (2001). Staphylococcal pore-forming toxins. Curr. Top. Microbiol. Immunol. 257, 53–83. doi: 10.1007/978-3-642-56508-3_4
Rasigade, J. P., Trouillet-Assant, S., Ferry, T., Diep, B. A., Sapin, A., Lhoste, Y., et al. (2013). PSMs of hypervirulent Staphylococcus aureus act as intracellular toxins that kill infected osteoblasts. PLoS ONE 8:e63176. doi: 10.1371/journal.pone.0063176
Reyes-Robles, T., Alonzo, F. 3rd., Kozhaya, L., Lacy, D. B., Unutmaz, D., and Torres, V. J. (2013). Staphylococcus aureus leukotoxin ED targets the chemokine receptors CXCR1 and CXCR2 to kill leukocytes and promote infection. Cell Host Microbe 14, 453–459. doi: 10.1016/j.chom.2013.09.005
Riegel, P., Jesel-Morel, L., Laventie, B., Boisset, S., Vandenesch, F., and Prevost, G. (2011). Coagulase-positive Staphylococcus pseudintermedius from animals causing human endocarditis. Int. J. Med. Microbiol. 301, 237–239. doi: 10.1016/j.ijmm.2010.09.001
Robb, A. R., Wright, E. D., Foster, A. M. E., Walker, R., and Malone, C. (2017). Skin infection caused by a novel strain of Staphylococcus pseudintermedius in a Siberian husky dog owner. JMM Case Rep. 4:jmmcr005087. doi: 10.1099/jmmcr.0.005087
Rubin, J. E., and Chirino-Trejo, M. (2011). Prevalence, sites of colonization, and antimicrobial resistance among Staphylococcus pseudintermedius isolated from healthy dogs in Saskatoon, Canada. J. Vet. Diagn Invest. 23, 351–354. doi: 10.1177/104063871102300227
Silva, M. B., Ferreira, F. A., Garcia, L. N., Silva-Carvalho, M. C., Botelho, L. A., Figueiredo, A. M., et al. (2015). An evaluation of matrix-assisted laser desorption ionization time-of-flight mass spectrometry for the identification of Staphylococcus pseudintermedius isolates from canine infections. J. Vet. Diagn. Invest. 27, 231–235. doi: 10.1177/1040638715573297
Somayaji, R., Priyantha, M. A., Rubin, J. E., and Church, D. (2016). Human infections due to Staphylococcus pseudintermedius, an emerging zoonosis of canine origin: report of 24 cases. Diagn. Microbiol. Infect. Dis. 85, 471–476. doi: 10.1016/j.diagmicrobio.2016.05.008
Spaan, A. N., Henry, T., van Rooijen, W. J., Perret, M., Badiou, C., Aerts, P. C., et al. (2013). The staphylococcal toxin Panton-Valentine Leukocidin targets human C5a receptors. Cell Host Microbe 13, 584–594. doi: 10.1016/j.chom.2013.04.006
Spaan, A. N., van Strijp, J. A. G., and Torres, V. J. (2017). Leukocidins: staphylococcal bi-component pore-forming toxins find their receptors. Nat. Rev. Microbiol. 15, 435–447. doi: 10.1038/nrmicro.2017.27
Starlander, G., Borjesson, S., Gronlund-Andersson, U., Tellgren-Roth, C., and Melhus, A. (2014). Cluster of infections caused by methicillin-resistant Staphylococcus pseudintermedius in humans in a tertiary hospital. J. Clin. Microbiol. 52, 3118–3120. doi: 10.1128/JCM.00703-14
Stegmann, R., Burnens, A., Maranta, C. A., and Perreten, V. (2010). Human infection associated with methicillin-resistant Staphylococcus pseudintermedius ST71. J. Antimicrob. Chemother. 65, 2047–2048. doi: 10.1093/jac/dkq241
Surewaard, B. G., Nijland, R., Spaan, A. N., Kruijtzer, J. A., de Haas, C. J., and van Strijp, J. A. (2012). Inactivation of staphylococcal phenol soluble modulins by serum lipoprotein particles. PLoS Pathog. 8:e1002606. doi: 10.1371/journal.ppat.1002606
Trouillet-Assant, S., Flammier, S., Sapin, A., Dupieux, C., Dumitrescu, O., Tristan, A., et al. (2015). Mupirocin resistance in isolates of Staphylococcus spp. from nasal swabs in a tertiary hospital in France. J. Clin. Microbiol. 53, 2713–2715. doi: 10.1128/JCM.00274-15
Tsompanidou, E., Denham, E. L., Becher, D., de Jong, A., Buist, G., van Oosten, M., et al. (2013). Distinct roles of phenol-soluble modulins in spreading of Staphylococcus aureus on wet surfaces. Appl. Environ. Microbiol. 79, 886–895. doi: 10.1128/AEM.03157-12
Van Hoovels, L., Vankeerberghen, A., Boel, A., Van Vaerenbergh, K., and De Beenhouwer, H. (2006). First case of Staphylococcus pseudintermedius infection in a human. J. Clin. Microbiol. 44, 4609–4612. doi: 10.1128/JCM.01308-06
Veldkamp, K. E., Heezius, H. C., Verhoef, J., van Strijp, J. A., and van Kessel, K. P. (2000). Modulation of neutrophil chemokine receptors by Staphylococcus aureus supernate. Infect. Immun. 68, 5908–5913. doi: 10.1128/IAI.68.10.5908-5913.2000
Wang, R., Braughton, K. R., Kretschmer, D., Bach, T. H., Queck, S. Y., Li, M., et al. (2007). Identification of novel cytolytic peptides as key virulence determinants for community-associated MRSA, (1510). Nat. Med. 13:4. doi: 10.1038/nm1656
Wang, R., Khan, B. A., Cheung, G. Y., Bach, T. H., Jameson-Lee, M., Kong, K. F., et al. (2011). Staphylococcus epidermidis surfactant peptides promote biofilm maturation and dissemination of biofilm-associated infection in mice. J. Clin. Invest. 121, 238–248. doi: 10.1172/JCI42520
Keywords: Staphylococcus pseudintermedius, pore-forming toxins, phenol-soluble modulins, leukotoxin Luk-I, CXCR2 receptor, cytotoxicity, non-professional phagocytic cells, polymorphonuclear neutrophils
Citation: Maali Y, Badiou C, Martins-Simões P, Hodille E, Bes M, Vandenesch F, Lina G, Diot A, Laurent F and Trouillet-Assant S (2018) Understanding the Virulence of Staphylococcus pseudintermedius: A Major Role of Pore-Forming Toxins. Front. Cell. Infect. Microbiol. 8:221. doi: 10.3389/fcimb.2018.00221
Received: 16 April 2018; Accepted: 11 June 2018;
Published: 28 June 2018.
Edited by:
Yinduo Ji, University of Minnesota Twin Cities, United StatesReviewed by:
Michael Otto, National Institute of Allergy and Infectious Diseases (NIAID), United StatesCopyright © 2018 Maali, Badiou, Martins-Simões, Hodille, Bes, Vandenesch, Lina, Diot, Laurent and Trouillet-Assant. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frederic Laurent, ZnJlZGVyaWMubGF1cmVudEB1bml2LWx5b24xLmZy
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.