 Emanuele Marzetti1,2
Emanuele Marzetti1,2 Flora Guerra3
Flora Guerra3 Riccardo Calvani1*
Riccardo Calvani1* Federico Marini4
Federico Marini4 Alessandra Biancolillo5Jacopo Gervasoni1,2Aniello Primiano2
Alessandra Biancolillo5Jacopo Gervasoni1,2Aniello Primiano2 Hélio José Coelho-Júnior2
Hélio José Coelho-Júnior2 Francesco Landi1,2Roberto Bernabei1,2
Francesco Landi1,2Roberto Bernabei1,2 Cecilia Bucci3
Cecilia Bucci3 Anna Picca1
Anna Picca1- 1Fondazione Policlinico Universitario “Agostino Gemelli” IRCCS, Rome, Italy
- 2Università Cattolica del Sacro Cuore, Rome, Italy
- 3Department of Biological and Environmental Sciences and Technologies, Università del Salento, Lecce, Italy
- 4Department of Chemistry, Sapienza Università di Roma, Rome, Italy
- 5Department of Physical and Chemical Sciences, Università degli Studi dell'Aquila, L'Aquila, Italy
Physical frailty and sarcopenia (PF&S) is a prototypical geriatric condition characterized by reduced physical function and low muscle mass. The multifaceted pathophysiology of this condition recapitulates all hallmarks of aging making the identification of specific biomarkers challenging. In the present study, we explored the relationship among three processes that are thought to be involved in PF&S (i.e., systemic inflammation, amino acid dysmetabolism, and mitochondrial dysfunction). We took advantage of the well-characterized cohort of older adults recruited in the “BIOmarkers associated with Sarcopenia and Physical frailty in EldeRly pErsons” (BIOSPHERE) study to preliminarily combine in a multi-platform analytical approach inflammatory biomolecules, amino acids and derivatives, and mitochondrial-derived vesicle (MDV) cargo molecules to evaluate their performance as possible biomarkers for PF&S. Eleven older adults aged 70 years and older with PF&S and 10 non-sarcopenic non-frail controls were included in the analysis based on the availability of the three categories of biomolecules. A sequential and orthogonalized covariance selection—linear discriminant analysis (SO-CovSel–LDA) approach was used for biomarkers selection. Of the 75 analytes assayed, 16 had concentrations below the detection limit. Within the remaining 59 biomolecules, So-CovSel–LDA selected a set comprising two amino acids (phosphoethanolamine and tryptophan), two cytokines (interleukin 1 receptor antagonist and macrophage inflammatory protein 1β), and MDV-derived nicotinamide adenine dinucleotide reduced form:ubiquinone oxidoreductase subunit S3 as the best predictors for discriminating older people with and without PF&S. The evaluation of these biomarkers in larger cohorts and their changes over time or in response to interventions may unveil specific pathogenetic pathways of PF&S and identify new biological targets for drug development.
Introduction
Sarcopenia is the progressive decline in muscle mass and strength that accompanies aging (Marzetti et al., 2017). This condition exposes older adults to a high risk of negative health-related events, including disability, loss of independence, institutionalization, and death (Marzetti et al., 2017). The public health relevance of sarcopenia is widely recognized and so is the need for effective preventive and therapeutic interventions (Beaudart et al., 2014). Yet, the heterogeneous phenotypic presentation of sarcopenia, the insufficient understanding of its pathophysiology, and the frequent superimposition of other age-related conditions have hampered the study of sarcopenia as a standalone phenomenon (Calvani et al., 2018a). This impasse is also reflected by the lack of a unique operational definition of sarcopenia (Landi et al., 2018) and clinically meaningful biomarkers (Calvani et al., 2017). In this scenario, the recently defined “physical frailty and sarcopenia” (PF&S) syndrome has marked a major step forward for the clinical and regulatory recognition of the condition (Cesari et al., 2017).
When digging into the pathways and processes involved in PF&S pathophysiology, several factors spanning from muscle-specific mitochondrial dysfunction to systemic changes (e.g., inflammation, amino acid dysmetabolism) have been pinpointed (Marzetti et al., 2016, 2019; Picca et al., 2017a; Calvani et al., 2018b). Whether these processes share common roots and how cell-based alterations spread and are sensed at the systemic level are presently unknown. Small extracellular vesicle (sEVs) of mitochondrial origin, termed mitochondrial-derived vesicles (MDVs), have recently been proposed as possible shuttles across biological systems (Picca et al., 2020a). However, little is known about their complex regulatory network.
The pathophysiology of PF&S recapitulates all hallmarks of aging (López-Otín et al., 2013; Sierra, 2016; Justice et al., 2018). Hence, PF&S is considered to be a prototypical geroscience conditions for which a strong interdependence and non-linear relationships between biomarkers may be envisioned (Cohen et al., 2018). In such a scenario, the analysis of single pathways enlightening discrete aspects of the condition and, thus, setting aside its multifaceted nature, might neglect relevant information (Cohen et al., 2018; Justice et al., 2018). This limitation could be overcome through the adoption of multivariate analytical strategies that enable the exploitation of more comprehensive, multi-platform datasets (Calvani et al., 2015). In the present preliminary study, we took advantage of the well-characterized cohort of older adults recruited in the “BIOmarkers associated with Sarcopenia and Physical frailty in EldeRly pErsons” (BIOSPHERE) study (Calvani et al., 2018b,c; Marzetti et al., 2019; Picca et al., 2019a, 2020a) to simultaneously analyze biomediators pertaining to three different domains: inflammation, amino acid metabolism, and mitochondrial quality control (MQC). The availability of systemic inflammatory and metabolic data from this cohort (Calvani et al., 2018b; Marzetti et al., 2019) and their complementation with the analysis of circulating MDVs (Picca et al., 2020a) provided a composite dataset to explore the relationship among systemic inflammation, metabolic characteristics, and MDV trafficking in PF&S. Data analysis was performed through sequential and orthogonalized covariance selection coupled with linear discriminant analysis (SO-CovSel–LDA), an innovative analytical strategy that is particularly suited for dealing with multi-block datasets (i.e., experimental settings in which variables are assayed using different platforms and/or at different time points) (Biancolillo et al., 2020). SO-CovSel–LDA enabled selecting the variables of interest for PF&S from a large number of highly correlated candidate biomarkers. The evaluation of these biomarkers in larger cohorts and their changes over time or in response to interventions may unveil specific pathophysiological pathways of PF&S and identify biological targets for drug development.
Materials and Methods
Participants
Participants of the present study were community-dwellers aged 70+ with PF&S and non-sarcopenic non-frail (nonPF&S) controls from the BIOSPHERE cohort (Calvani et al., 2018c). BIOSPHERE is a cross-sectional study conceived for selecting and validating a panel of candidate biomarkers for PF&S through multivariate statistical modeling (Calvani et al., 2018b,c; Picca et al., 2019a).
To diagnose PF&S, the operational definition developed in the “Sarcopenia and Physical fRailty IN older people: multi-componenT Treatment strategies” (SPRINTT) project (Marzetti et al., 2015, 2018) was applied: (a) physical frailty, based on a summary score on the Short Physical Performance Battery (SPPB) (Guralnik et al., 1994) between 3 and 9; (b) low appendicular muscle mass (aLM), according to the cut-points of the Foundation for the National Institutes of Health Sarcopenia Project (Studenski et al., 2014); and (c) absence of major mobility disability (i.e., inability to complete a 400-m walk test) (Newman et al., 2006). Data analysis for the present investigation was conducted in a convenience sample of 21 participants (11 older adults with PF&S and 10 nonPF&S controls) for whom complete data were available for inflammatory and metabolic mediators and MDV characterization.
The Ethics Committee of the Università Cattolica del Sacro Cuore (Rome, Italy; protocol number BIOSPHERE: 8498/15) approved the study protocol. All procedures were conducted in compliance to the ethical standards laid down in the 1964 Declaration of Helsinki and its later amendments. Study procedures and criteria for participant selection were described thoroughly elsewhere (Calvani et al., 2018c). A written inform consent form was signed by all participants prior to enrolment.
Assessment of Appendicular Lean Mass by Dual X-Ray Absorptiometry
Appendicular lean mass was quantified through whole-body dual X-ray absorptiometry scans on a Hologic Discovery A densitometer (Hologic, Inc., Bedford, MA, USA) according to the manufacturer's directions. Criteria for low aLM were as follows: (a) aLM to body mass index (BMI) ratio (aLMBMI) <0.789 and <0.512 in men and women, respectively; or (b) crude aLM <19.75 kg in men and <15.02 kg in women when the aLMBMI criterion was not met (Studenski et al., 2014).
Collection and Processing of Blood Samples
Blood samples were obtained in the morning after overnight fasting by venipuncture of the median cubital vein, using BD Vacutainer® commercial tubes (Becton, Dickinson and Co., Franklin Lakes, NJ, USA). One tube was delivered to the centralized diagnostic laboratory of the Fondazione Policlinico Universitario “Agostino Gemelli” IRCCS (Rome, Italy) for standard blood biochemistry. The remaining tubes were processed for serum separation in the Biogerontology lab of the Università Cattolica del Sacro Cuore. After 30 min of blood clotting at room temperature, samples were centrifuged at 1,000 × g for 15 min at 4°C. The upper clear fraction (serum) was recovered in 0.5-mL aliquots and stored at −80°C until analysis.
Purification of Small Extracellular Vesicles From Serum
Small EVs were purified from serum through differential centrifugation and were quantified as previously reported (Picca et al., 2018a, 2019b, 2020a). In brief, serum samples were diluted with equal volumes of phosphate-buffered saline (PBS) and centrifuged at 2,000 × g at 4°C for 30 min. Supernatants were collected and centrifuged at 12,000 × g at 4°C for 45 min to remove apoptotic bodies, mitochondrial fragments, cell debris, and vesicles larger than 200 nm. Supernatants were subsequently ultracentrifuged at 110,000 × g at 4°C for 2 h. Pellets were recovered and resuspended in PBS, filtered through a 0.22-μm filter and ultracentrifuged at 110,000 × g at 4°C for 70 min to eliminate contaminant proteins. Pellets enriched in purified sEVs were finally resuspended in 100 μL of PBS. To quantify sEVs, total protein concentration was measured using the Bradford assay (Théry et al., 2006).
Measurement of Inflammatory, Metabolic, and Mitochondrial Markers
Serum samples were assayed for a panel of 75 candidate biomarkers (Table 1). The panel was designed based on previous studies in older adults and their involvement in pathways and processes relevant to PF&S pathophysiology (i.e., inflammation, amino acid metabolism, and mitochondrial dysfunction) (Calvani et al., 2018b, 2020a; Marzetti et al., 2019; Picca et al., 2019a, 2020b).

Table 1. Composition of the biomarker panel.
Twenty-seven inflammatory mediators including cytokines, chemokines, and growth factors were assayed using the Bio-Plex Pro™ Human Cytokine 27-plex Assay kit (#M500KCAF0Y, Bio-Rad Laboratories Inc., Hercules, CA, USA) on a Bio-Plex® System with Luminex xMAP® Technology (Bio-Rad Laboratories), as described elsewhere (Ponziani et al., 2018, 2019; Marzetti et al., 2019; Picca et al., 2019a; Addolorato et al., 2020). Serum levels of C-reactive protein (CRP), myeloperoxidase (MPO), fibroblast growth factor (FGF) 21, and brain-derived neurotrophic factor (BDNF) were measured by using commercially available kits on an ELLA™ automated immunoassay system (Bio-Techne, San Jose, CA, USA). The concentration of 37 amino acids and derivatives was determined by ultraperformance liquid chromatography/mass spectrometry (UPLC/MS) as described previously (Calvani et al., 2018b).
Protein levels of tetraspanin CD63 and selected mitochondrial markers were measured by Western immunoblot analysis in purified sEVs, as detailed elsewhere (Picca et al., 2020a). According to the guidelines of the International Society of Extracellular Vesicles (Théry et al., 2018), sEV purity was ascertained by verifying the presence of the cytosolic protein flotilin (positive control) and the absence of the non-sEV component heterogeneous nuclear ribonucleoprotein A1 (HNRNPA1, negative control). Scanning electron microscopy analyses were performed to confirm enrichment in sEVs (Théry et al., 2018).
Statistical Analysis
Descriptive Statistics
Descriptive statistics were run on all data. The normal distribution of data was ascertained via the Kolmogorov–Smirnov test. Differences in demographic, anthropometric, clinical, and biological parameters between participants with and without PF&S were assessed via t-test statistics or the Mann–Whitney U-test, as appropriate, for continuous variables. Chi-squared χ2 or Fisher exact tests were applied for categorical variables. All tests were two-sided, with statistical significance set at p < 0.05. Analyses were performed using the GraphPrism 5.03 software (GraphPad Software, Inc., San Diego, CA, USA).
Sequential and Orthogonalized Covariance Selection – Linear Discriminant Analysis
In order to identify relationships among different sets of data and to identify putative markers of PF&S, a recently proposed multi-block classification approach, called SO-CovSel–LDA (Biancolillo et al., 2020), was adopted. SO-CovSel–LDA is a highly efficient multi-block classification strategy, which combines a very parsimonious variable selection algorithm to be applied on each individual block (CovSel) (Roger et al., 2011) with the sequential inclusion of data matrices, after orthogonalization with respect to the previously selected variables. This procedure reduces redundancies among the blocks and allows a clearer interpretation of results.
Here, we begin with describing the CovSel variable selection algorithm for a single block of predictors X to make its generalization to the case of multiple blocks easier to follow. CovSel allows selecting the minimum number of variables that provide an accurate regression model between the X matrix and a response Y. Variables are progressively selected from the X-block as those having maximum covariance with Y. Parsimony is achieved as any successive predictor is selected after both X and Y are orthogonalized with respect to the previously chosen variables, so to bring as much new information as possible. SO-CovSel is a generalization of this procedure to a multi-block case, enriched by the concept of sequential inclusion of the blocks after orthogonalization which is borrowed from methods such as SO partial least squares (SO-PLS) regression (Biancolillo and Næs, 2019; Biancolillo et al., 2019). In the case of three blocks of predictors, like in the present study (i.e., inflammatory markers, amino acids, and MDV cargo molecules), the SO-CovSel algorithm proceeds as follows. Variables are selected from the first block (amino acids and derivatives) using CovSel, then both the second block (inflammatory markers) X2 and the Y are orthogonalized with respect to the variables selected from the first block (X1,sel):
where I is the identity matrix and the superscript T indicates matrix transposition. CovSel is then applied to the matrix X2,orth to select variables having maximum covariance with Yorth; the selected variables are subsequently collected in the matrix X2,sel. The third block (MDV markers) and the Y are orthogonalized with respect to the variables selected from the first two blocks, similar to what described in equations (1) and (2). CovSel is applied to the orthogonalized third block X3,orth to select variables (X3,sel) with maximum covariance with the orthogonalized response. Finally, an overall regression model is built between the selected variables from the three blocks and the Y:
where is the predicted response and B1, B2, and B3 are the regression coefficients.
SO-CovSel–LDA is the classification analog of the SO-CovSel regression algorithm. As such, it requires the class information to be encoded in a binary-coded response vector y. The response vector will have the value 1 for PF&S participants and 0 for nonPF&S controls. A SO-CovSel model is built between the different blocks of predictors and the y as described above. Eventually, LDA is applied to the predicted response to accomplish sample classification.
To unbiasedly validate the results, a repeated double cross-validation (rDCV) procedure was used (Smit et al., 2007; Biancolillo et al., 2019). DCV consists of two loops of cross-validation nested into one another: the inner loop is used for model selection and the outer loop for external validation (i.e., contains samples that were not use at any stage of model building or optimization). Since DCV implies splitting the samples into the different cross-validation groups, to avoid the outcomes depending on a particular splitting scheme, the whole procedure is repeated a certain number of times (50 in the present study). In the context of biomarker discovery, the use of rDCV has the advantage that many different models are calculated (as many as the product of the number of DCV runs times the number of splits in the inner loop), which enables evaluating how consistently variables are selected by SO-CovSel–LDA and how robust candidate markers are. All calculations were carried out using in-house written functions running under Matlab environment (R2015b, The Mathworks, Natick, MA) and freely downloadable at: https://www.chem.uniroma1.it/romechemometrics/research/algorithms/so-covsel/.
Results
Characteristics of the Study Participants
Data from 21 BIOSPHERE participants, 11 older adults with PF&S (mean age 77.7 ± 5.4 years; 73.0% women) and 10 nonPF&S controls (mean age 73.9 ± 2.7 years; 50.0% women) were analyzed in the present study. Demographic, functional, anthropometric, and clinical characteristics of the included participants were comparable to those of the whole BIOSPHERE cohort (Marzetti et al., 2019).
As shown in Table 2, no differences were observed in sex distribution, BMI, or number of comorbid conditions and medications between PF&S and controls participants. The latter group was slightly younger, but the age difference did not reach statistical significance. Serum albumin, total serum protein concentration, and the neutrophil count were comparable between groups. On the other hand, the lymphocyte count was lower and the neutrophil-to-lymphocyte ratio was higher in the PF&S group. As per the inclusion criteria adopted, participants with PF&S had lower SPPB scores and smaller aLM either crude or adjusted by BMI compared with nonPF&S controls.
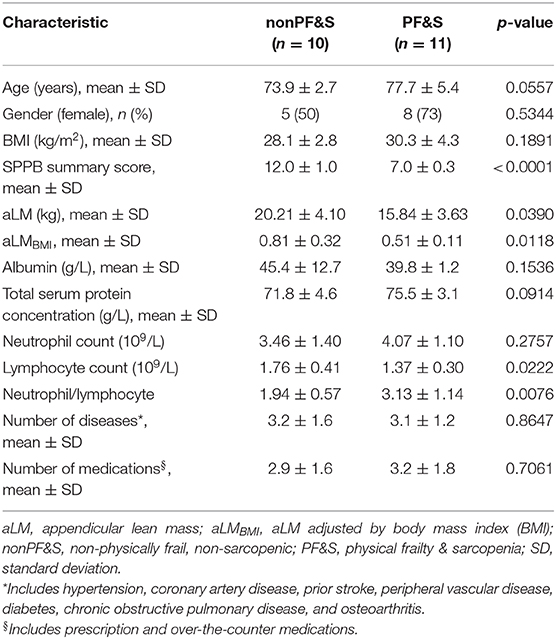
Table 2. Participant characteristics according to the presence of physical frailty & sarcopenia.
Biomarker Selection Through SO-CovSel–LDA Analysis
Serum levels of 75 inflammatory cytokines, growth factors, neurogenesis and neural plasticity mediators, amino acids and derivatives, and MDV cargo molecules were measured through multiple analytical platforms. Of the assayed biomolecules, concentrations of anserine, carnosine, cystathionine, γ-aminobutyric acid, phosphoserine, interleukin (IL) 2, IL5, IL7, IL10, IL13, IL15, granulocyte colony-stimulating factor, vascular endothelial growth factor, mitochondrial cytochrome C oxidase subunit I, nicotinamide adenine dinucleotide reduced form (NADH):ubiquinone oxidoreductase subunit B8, and ubiquinol-cytochrome C reductase core protein 2 were below the detection limit. Serum concentrations of the assayed biomolecules are shown in Supplementary Table 1. SO-CovSel–LDA models were built using a multi-matrix dataset containing 59 analytes. Serum concentrations of candidate biomarkers were organized into three matrices according to the analytical approach adopted for their determination and the biological domain of pertinence (Table 1). Prior to data analysis, blocks were individually pretreated by autoscaling through subtracting from each variable its mean and dividing the result by its standard deviation. Afterwards, SO-CovSel–LDA models were built and validated via 50 runs of rDCV with 21 cancelation groups in the outer loop and five in the inner loop. The optimal order of blocks and the optimal number of variables from each block were selected as those returning the smallest error in the inner cross-validation loop. Since a total of 50 × 21 (i.e., 1,050) different models were built, the procedure allowed calculating the confidence intervals for the predictive ability and the consistency of selection of the optimal number of predictors as well as revealing their identity.
The predictive ability was evaluated on the samples of the outer rDCV loop, since they were completely external of the models used for their prediction, thereby providing more unbiased estimates of the discriminant capacity. Our analytical approach was able to correctly classify 87.5 ± 7.3% of PF&S participants and 83.6 ± 9.2% of nonPF&S controls, corresponding to an overall classification accuracy of 85.6 ± 5.0%. Permutation tests with 1,000 randomizations, which describe the distribution of figures of merit under the null hypothesis, showed that the discriminant ability of the model was statistically significant (p < 0.001).
Results appeared to be highly consistent across rDCV runs. Indeed, in all of the 1,050 models calculated, the best order of the blocks was found to be (1) amino acids and derivatives, (2) inflammatory biomolecules, and (3) MDV markers. In the large majority of iterations, the optimal model complexity was found to be 2, 2, and 1 variables, respectively. Figure 1 illustrates the frequency of selection of discriminant markers across the 1,050 models calculated during the rDCV procedure. The figure shows that the optimal model complexity was consistently found to be 2-2-1, and that the same biomolecules—i.e., phosphoethanolamine, tryptophan, IL1 receptor antagonist (IL1-ra), macrophage inflammatory protein 1β (MIP-1β), and NDUF subunit S3 (NDUFS3)—were selected in almost all iterations, thus confirming the robustness of the proposed biomarkers in our sample.
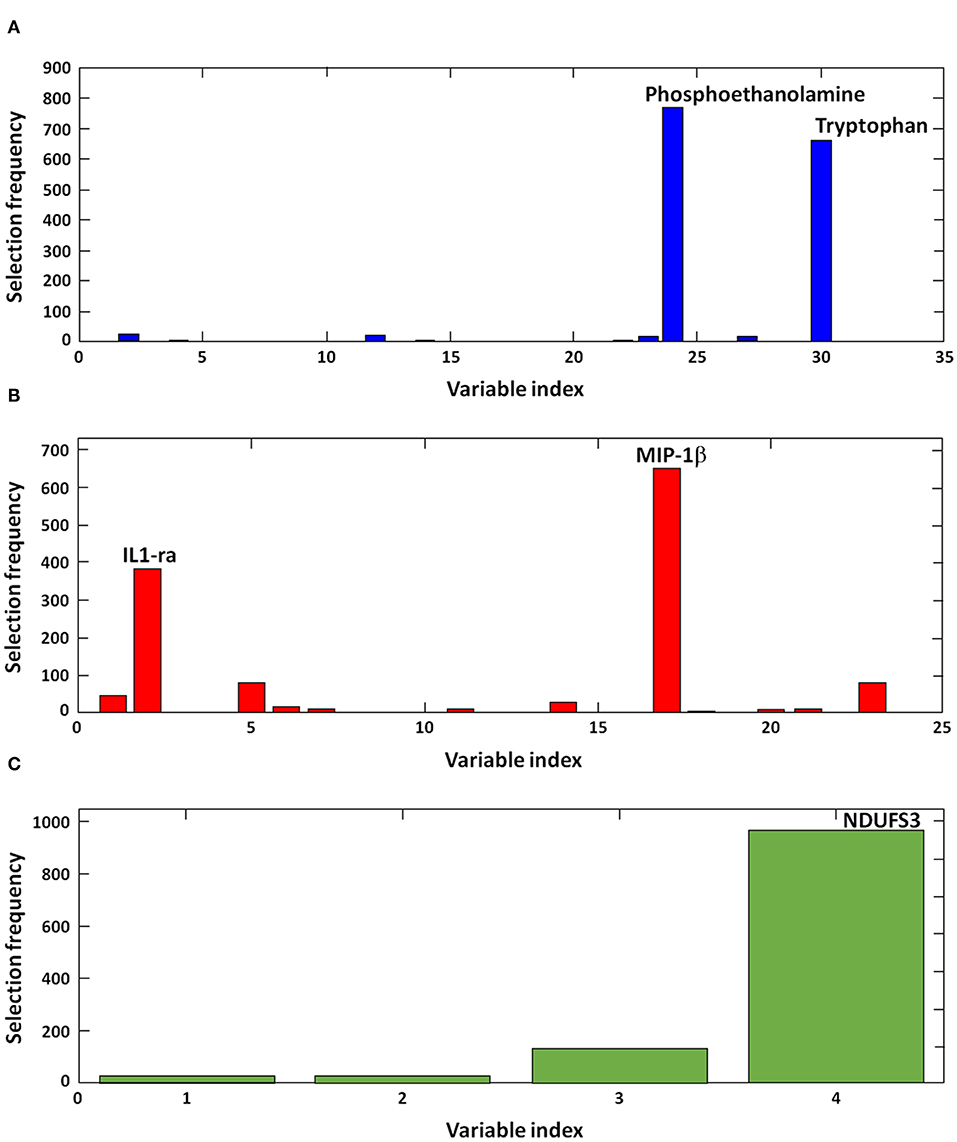
Figure 1. Discriminant biomolecules selected by sequential and orthogonalized covariance selection coupled with linear discriminant analysis (SO-CovSel–LDA). Candidate biomarkers are presented according to the order in which they respective data matrices were entered into the model: (A) amino acids and derivatives, (B) inflammatory biomolecules, and (C) mitochondrial-derived vesicle cargo molecules. IL1-ra, interleukin 1 receptor agonist; MIP-1β, macrophage inflammatory protein 1β; NDFUS3, nicotinamide adenine dinucleotide reduced form (NADH), ubiquinone oxidoreductase subunit S3.
Discussion
According to the geroscience paradigm, the roots of most chronic diseases reside in perturbations of a discrete set of biological mechanisms, collectively termed hallmarks of aging (López-Otín et al., 2013). The pathophysiology of sarcopenia involves all major biological pillars of aging and is, therefore, envisioned as a prototypical geroscience condition (Calvani et al., 2020a). In particular, derangements of skeletal myocyte quality control mechanisms are thought to play a relevant role in the development and progression of age-related muscle wasting (Iqbal et al., 2013). Indeed, altered recycling of damaged cell components and organelles via autophagy, defective mitochondrial proteostasis and dynamics, and impaired mitochondriogenesis have been described in muscles of older adults with PF&S (Picca et al., 2018a). Systemic signatures of PF&S, including specific inflammatory and amino acid profiles, have also been identified (Calvani et al., 2018b, 2020a; Marzetti et al., 2019).
In the present study, we applied an innovative SO-CovSel–LDA analytical approach to explore the relationship among systemic inflammation, metabolic derangements, and circulating MDVs in PF&S in order to provide hints on the underlying pathogenic mechanisms. Among all assayed molecules, SO-CovSel–LDA selected a panel comprising two amino acids (i.e., phosphoethanolamine and tryptophan), two cytokines (i.e., IL1-ra and MIP-1β), and NDUFS3, a subunit of complex I of the mitochondrial electron transport chain (ETC), as the best predictors to discriminate older adults with and without PF&S.
The identification of phosphoethanolamine within the discriminant metabolites for the classification of PF&S is particularly relevant. Phosphoethanolamine is an ethanolamine derivate produced as an intermediate of the CDP-ethanolamine pathway involved in the metabolism of glycerophospholipid and biological membrane turnover (Patel and Witt, 2017; van der Veen et al., 2017). Disruption of CDP-ethanolamine pathway has been associated with mitochondrial dyshomeostasis in mouse models of muscle atrophy (Selathurai et al., 2019) and insulin resistance (Funai et al., 2016). Phosphoethanolamine has also been found to mediate mitochondrial membrane fusion and curvatures and, in combination with ethanolamine, to promote autophagy and longevity (Rockenfeller et al., 2015). The difference in phosphoethanolamine serum levels between older adults with and without PF&S might reflect impairment of autophagy in the setting of muscle atrophy (Iqbal et al., 2013; Marzetti et al., 2016). This hypothesis is in keeping with the higher secretion of sEVs previously described in older adults with PF&S (Picca et al., 2020a), which could be interpreted as an attempt to cope with deficient MQC processes (Soubannier et al., 2012; Picca et al., 2020b). Noticeably, derangements in the MQC machinery were documented in intraoperative muscle biopsies obtained from old hip-fractured patients with sarcopenia (Marzetti et al., 2016). Finally, phosphoethanolamine was also found among the mediators possibly involved in the disabling cascade in frail older persons with type 2 diabetes mellitus (T2DM) (Calvani et al., 2020b).
The amino acidic profile of participants with PF&S also included the aromatic essential amino acid tryptophan. The latter regulates several activities within the body, including growth, mood, behavior, and immune responses (Le Floc'h et al., 2011). Its metabolism is mediated by the tryptophan-kynurenine and the tryptophan-methoxyndole pathways, leading to the production of physiologically relevant bioactive compounds, such as NAD, serotonin, and melatonin (Le Floc'h et al., 2011). Changes in tryptophan circulating levels have been associated with low muscle quality (Moaddel et al., 2016), insulin resistance, and frailty in older adults with T2DM (Chen et al., 2016; Marcos-Pérez et al., 2017; Calvani et al., 2020b).
As per the inflammatory fingerprint of PF&S, IL1-ra and MIP-1β were selected by SO-CovSel–LDA as the most relevant mediators. A frailty “cytokinome” in older adults with PF&S composed of a “core” inflammatory profile with gender-specific signatures was previously described by our group (Marzetti et al., 2019). In a later study, SO-CovSel allowed restricting to only MPO and platelet-derived growth factor BB the mediators describing the contribution of inflammation to PF&S (Calvani et al., 2020a). In the same study, gender-specific models selected MIP-1β as one of the most relevant biomarkers for the discrimination between PF&S and nonPF&S participants (Calvani et al., 2020a). MIP-1β is a chemokine that regulates myoblast response to muscle injury and promotes leucocyte recruitment at the site of muscle damage (Yahiaoui et al., 2008). The presence of MIP-1β among discriminant analytes for PF&S may be interpreted as a compensatory action to impaired muscle regenerative capacity (Calvani et al., 2020a). Inflamm-aging, the chronic low-grade inflammation that develops during aging, has been involved in the pathogenesis of sarcopenia and physical disability (Wilson et al., 2017; Franceschi et al., 2018; Furman et al., 2019). In this context can be framed the selection of IL1-ra among the discriminant biomolecules associated with PF&S. IL1-ra is a natural negative modulator of IL1α- and IL1β-mediated inflammatory response and was found to be overexpressed in muscles of older adults (Przybyla et al., 2006). The inflammatory milieu of PF&S is also reflected by the higher neutrophil-to-lymphocyte ratio, an easily accessible indicator of inflammation.
Finally, the MDV cargo molecule NDUFS3 was selected among the discriminant mediators for PF&S. NDUFS3 is a nuclear-encoded subunit of complex I of the mitochondrial ETC. Mutations of NDUFS3 are associated with deficiency in mitochondrial respiration and myopathies (Bénit et al., 2004; Pereira et al., 2020). Conversely, NDUFS3 gene replacement rescued muscle structure and mitochondrial in a mouse model of mitochondrial myopathy (Pereira et al., 2020). Mitochondrial dysfunction and insufficient MDV-mediated quality control have also been hypothesized to contribute to PF&S (Picca et al., 2020a). This idea is in keeping with the altered expression of key MQC proteins described in muscles of old hip-fractured patients with sarcopenia (Marzetti et al., 2016).
Taken as a whole, our findings provide preliminary, yet novel insights into the relationship among metabolic changes, inflamm-aging, and mitochondrial dyshomeostasis in PF&S. In particular, the retrieval of MDVs in serum of older adults with PF&S allows placing this process in the context of an innate immune response as part of the “danger theory” of inflammation (Zhang et al., 2010). According to this view, MDVs may function as antigen-presenting vesicles carrying misplaced noxious material. Similar to damage-associated molecular patterns (DAMPs) released from injured cells, the MDV cargo can trigger caspase-1 activation and the secretion of pro-inflammatory cytokines (Krysko et al., 2011). This inflammatory response would be mounted through the interactions of mitochondrial DAMPs with receptors/systems including Toll-like receptors, Nod-like receptor family pyrin domain containing 3 inflammasome, and cytosolic cyclic GMP–AMP synthase-stimulator of interferon genes DNA sensing system (Picca et al., 2017b). Impaired MQC in skeletal myocytes may therefore install a vicious circle favoring further mitochondrial damage and the propagation of sterile inflammation through DAMPs release (Picca et al., 2018b). Should this hypothesis be confirmed, the scavenging of circulating mitochondrial DAMPs might be exploited for the development of therapeutic interventions for PF&S.
Albeit presenting promising findings, our work has limitations that deserve discussion. First of all, the study was conducted in a small participant sample and results need to be validated in larger cohorts. Although participants were carefully selected and thoroughly characterized, the chance of unknown comorbidities affecting the results cannot be excluded. Also, the cross-sectional design of the study hampers establishing cause-effect or temporal relationships between the analyzed pathways and PF&S development. Finally, while the biomarker panel analyzed in the present study included a large number of biomolecules, the possibility exists that a more comprehensive appraisal of PF&S pathophysiology might be obtained through the analysis of larger sets of mediators.
Data Availability Statement
All datasets generated for this study are included in the article/Supplementary Material.
Ethics Statement
The studies involving human participants were reviewed and approved by Università Cattolica del Sacro Cuore, Rome, Italy. The participants provided their written informed consent to participate in this study.
Author Contributions
APi, CB, EM, FG, and RC: conceptualization. APi and FG: data curation. AB and FM: data analysis. APi, APr, FG, HC-J, and JG: methodology. APi, EM, and RC: writing—original draft preparation. CB, FG, and FL: writing—review and editing. FL and RB: supervision. CB and RB: funding acquisition. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by Innovative Medicine Initiative-Joint Undertaking (IMI-JU #115621), AIRC (Associazione Italiana per la Ricerca sul Cancro) Investigator grant 2016 #19068 to CB, Ministero dell'Istruzione, dell'Università e della Ricerca (MIUR) to Consorzio Interuniversitario Biotecnologie (DM 1049, 29/12/2018; CIB N. 112/19 to CB), intramural research grants from the Università Cattolica del Sacro Cuore (D3.2 2013, D3.2 2015, and D3.2 2020), and the nonprofit research foundation Centro Studi Achille e Linda Lorenzon.
Conflict of Interest
EM served as a consultant for Abbott, Biophytis, Nutricia, and Nestlè. RC served as a consultant for Abbott and Nutricia.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor declared a past co-authorship with several of the authors EM, RC, and AP.
The reviewers FS and EP declared a past collaboration with one of the authors EM to the handling editor.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2020.564417/full#supplementary-material
References
Addolorato, G., Ponziani, F. R., Dionisi, T., Mosoni, C., Vassallo, G. A., Sestito, L., et al. (2020). Gut microbiota compositional and functional fingerprint in patients with alcohol use disorder and alcohol-associated liver disease. Liver Int. 40, 878–88. doi: 10.1111/liv.14383
Beaudart, C., Rizzoli, R., Bruyère, O., Reginster, J. Y., and Biver, E. (2014). Sarcopenia: burden and challenges for public health. Arch Public Health 72:45. doi: 10.1186/2049-3258-72-45
Bénit, P., Slama, A., Cartault, F., Giurgea, I., Chretien, D., Lebon, S., et al. (2004). Mutant NDUFS3 subunit of mitochondrial complex I causes leigh syndrome. J. Med. Genet. 41, 14–17. doi: 10.1136/jmg.2003.014316
Biancolillo, A., Boqué, R., Cocchi, M., and Marini, F. (2019). “Data fusion strategies in food analysis,” in: Cocchi M, editor. Data Fusion Methodology and Application. Oxford: Elsevier Ltd. 271–310. doi: 10.1016/B978-0-444-63984-4.00010-7
Biancolillo, A., Marini, F., and Roger, J. M. (2020). SO-CovSel: a novel method for variable selection in a multiblock framework. J. Chemom. 34:e3120. doi: 10.1002/cem.3120
Biancolillo, A., and Næs, T. (2019). “The sequential and orthogonalized PLS regression for multiblock regression: theory, examples, and extensions,” in: Cocchi M, editor. Data Fusion Methodology and Application. Oxford: Elsevier Ltd. 157–77.
Calvani, R., Marini, F., Cesari, M., Tosato, M., Anker, S. D., von Haehling, S., et al. (2015). Biomarkers for physical frailty and sarcopenia: state of the science and future developments. J. Cachexia Sarcopenia Muscle 6, 278–286. doi: 10.1002/jcsm.12051
Calvani, R., Marini, F., Cesari, M., Tosato, M., Picca, A., Anker, S. D., et al. (2017). Biomarkers for physical frailty and sarcopenia. Aging Clin. Exp. Res. 29, 29–34. doi: 10.1007/s40520-016-0708-1
Calvani, R., Picca, A., Cesari, M., Tosato, M., Marini, F., Manes-Gravina, E., et al. (2018a). Biomarkers for sarcopenia: reductionism vs. complexity. Curr. Protein Pept. Sci. 19, 639–642. doi: 10.2174/1389203718666170516115422
Calvani, R., Picca, A., Marini, F., Biancolillo, A., Cesari, M., Pesce, V., et al. (2018c). The “BIOmarkers associated with Sarcopenia and PHysical frailty in EldeRly pErsons” (BIOSPHERE) study: rationale, design and methods. Eur. J. Intern. Med. 56, 19–25. doi: 10.1016/j.ejim.2018.05.001
Calvani, R., Picca, A., Marini, F., Biancolillo, A., Gervasoni, J., Persichilli, S., et al. (2018b). A distinct pattern of circulating amino acids characterizes older persons with physical frailty and sarcopenia: results from the BIOSPHERE study. Nutrients 10:1691. doi: 10.3390/nu10111691
Calvani, R., Picca, A., Marini, F., Biancolillo, A., Gervasoni, J., Persichilli, S., et al. (2020a). Identification of biomarkers for physical frailty and sarcopenia through a new multi-marker approach: results from the BIOSPHERE study. GeroScience. doi: 10.1007/s11357-020-00197-x
Calvani, R., Rodriguez-Mañas, L., Picca, A., Marini, F., Biancolillo, A., Laosa, O., et al. (2020b). Identification of a circulating amino acid signature in frail older persons with type 2 diabetes mellitus: results from the Metabofrail study. Nutrients 12:199. doi: 10.3390/nu12010199
Cesari, M., Landi, F., Calvani, R., Cherubini, A., Di Bari, M., Kortebein, P., et al. (2017). Rationale for a preliminary operational definition of physical frailty and sarcopenia in the SPRINTT trial. Aging Clin. Exp. Res. 29, 81–88. doi: 10.1007/s40520-016-0716-1
Chen, T., Zheng, X., Ma, X., Bao, Y., Ni, Y., Hu, C., et al. (2016). Tryptophan predicts the risk for future type 2 diabetes. PLoS ONE 11:e0162192. doi: 10.1371/journal.pone.0162192
Cohen, A. A., Legault, V., Fuellen, G., Fülöp, T., Fried, L. P., and Ferrucci, L. (2018). The risks of biomarker-based epidemiology: associations of circulating calcium levels with age, mortality, and frailty vary substantially across populations. Exp. Gerontol. 107:11–17. doi: 10.1016/j.exger.2017.07.011
Franceschi, C., Garagnani, P., Parini, P., Giuliani, C., and Santoro, A. (2018). Inflammaging: a new immune–metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 14, 576–590. doi: 10.1038/s41574-018-0059-4
Funai, K., Lodhi, I. J., Spears, L. D., Yin, L., Song, H., Klein, S., et al. (2016). Skeletal muscle phospholipid metabolism regulates insulin sensitivity and contractile function. Diabetes 65, 358–370. doi: 10.2337/db15-0659
Furman, D., Campisi, J., Verdin, E., Carrera-Bastos, P., Targ, S., Franceschi, C., et al. (2019). Chronic inflammation in the etiology of disease across the life span. Nat. Med. 25, 1822–1832. doi: 10.1038/s41591-019-0675-0
Guralnik, J. M., Simonsick, E. M., Ferrucci, L., Glynn, R. J., Berkman, L. F., Blazer, D. G., et al. (1994). A short physical performance battery assessing lower extremity function: association with self-reported disability and prediction of mortality and nursing home admission. J. Gerontol. 49, M85–M94. doi: 10.1093/geronj/49.2.M85
Iqbal, S., Ostojic, O., Singh, K., Joseph, A.-M., and Hood, D. A. (2013). Expression of mitochondrial fission and fusion regulatory proteins in skeletal muscle during chronic use and disuse. Muscle Nerve 48, 963–970. doi: 10.1002/mus.23838
Justice, J. N., Ferrucci, L., Newman, A. B., Aroda, V. R., Bahnson, J. L., Divers, J., et al. (2018). A framework for selection of blood-based biomarkers for geroscience-guided clinical trials: report from the TAME Biomarkers Workgroup. GeroScience 40, 419–436. doi: 10.1007/s11357-018-0042-y
Krysko, D. V., Agostinis, P., Krysko, O., Garg, A. D., Bachert, C., Lambrecht, B. N., et al. (2011). Emerging role DAMPs derived from mitochondria in inflammation. Trends Immunol. 32, 157–164. doi: 10.1016/j.it.2011.01.005
Landi, F., Calvani, R., Cesari, M., Tosato, M., Maria Martone, A., Ortolani, E., et al. (2018). Sarcopenia: an overview on current definitions, diagnosis and treatment. Curr. Protein Pept. Sci. 19, 633–638. doi: 10.2174/1389203718666170607113459
Le Floc'h, N., Otten, W., and Merlot, E. (2011). Tryptophan metabolism, from nutrition to potential therapeutic applications. Amino Acids 41, 1195–1205. doi: 10.1007/s00726-010-0752-7
López-Otín, C., Blasco, M. A., Partridge, L., Serrano, M., and Kroemer, G. (2013). The hallmarks of aging. Cell 153, 1194–1217. doi: 10.1016/j.cell.2013.05.039
Marcos-Pérez, D., Sánchez-Flores, M., Maseda, A., Lorenzo-López, L., Millán-Calenti, J. C., Strasser, B., et al. (2017). Frailty Status in older adults is related to alterations in indoleamine 2,3-dioxygenase 1 and guanosine triphosphate cyclohydrolase I enzymatic pathways. J. Am. Med. Dir. Assoc. 18, 1049–1057. doi: 10.1016/j.jamda.2017.06.021
Marzetti, E., Calvani, R., Landi, F., Hoogendijk, E., Fougère, B., Vellas, B., et al. (2015). Innovative medicines initiative: the SPRINT project. J. Frailty Aging 4, 207–208. doi: 10.14283/jfa.2015.69
Marzetti, E., Calvani, R., Lorenzi, M., Tanganelli, F., Picca, A., Bossola, M., et al. (2016). Association between myocyte quality control signaling and sarcopenia in old hip-fractured patients: results from the Sarcopenia in HIp FracTure (SHIFT) exploratory study. Exp. Gerontol. 80, 1–5. doi: 10.1016/j.exger.2016.04.003
Marzetti, E., Calvani, R., Tosato, M., Cesari, M., Di Bari, M., Cherubini, A., et al. (2017). Sarcopenia: an overview. Aging Clin. Exp. Res. 29, 11–17. doi: 10.1007/s40520-016-0704-5
Marzetti, E., Cesari, M., Calvani, R., Msihid, J., Tosato, M., Rodriguez-Mañas, L., et al. (2018). The “Sarcopenia and Physical fRailty IN older people: multi-componenT Treatment strategies” (SPRINTT) randomized controlled trial: case finding, screening and characteristics of eligible participants. Exp. Gerontol. 113, 48–57. doi: 10.1016/j.exger.2018.09.017
Marzetti, E., Picca, A., Marini, F., Biancolillo, A., Coelho-Junior, H. J., Gervasoni, J., et al. (2019). Inflammatory signatures in older persons with physical frailty and sarcopenia: the frailty “cytokinome” at its core. Exp. Gerontol. 122, 129–138. doi: 10.1016/j.exger.2019.04.019
Moaddel, R., Fabbri, E., Khadeer, M. A., Carlson, O. D., Gonzalez-Freire, M., Zhang, P., et al. (2016). Plasma biomarkers of poor muscle quality in older men and women from the Baltimore Longitudinal Study of Aging. J. Gerontol A Biol. Sci. Med. Sci. 71, 1266–1272. doi: 10.1093/gerona/glw046
Newman, A. B., Simonsick, E. M., Naydeck, B. L., Boudreau, R. M., Kritchevsky, S. B., Nevitt, M. C., et al. (2006). Association of long-distance corridor walk performance with mortality, cardiovascular disease, mobility limitation, and disability. J. Am. Med. Assoc. 295, 2018–2026. doi: 10.1001/jama.295.17.2018
Patel, D., and Witt, S. N. (2017). Ethanolamine and phosphatidylethanolamine: partners in health and disease. Oxid. Med. Cell. Longev. 2017:4829180. doi: 10.1155/2017/4829180
Pereira, C. V., Peralta, S., Arguello, T., Bacman, S. R., Diaz, F., and Moraes, C. T. (2020). Myopathy reversion in mice after restauration of mitochondrial complex I. EMBO Mol. Med. 12:e10674. doi: 10.15252/emmm.201910674
Picca, A., Beli, R., Calvani, R., Coelho-Júnior, H. J., Landi, F., Bernabei, R., et al. (2020a). Older adults with physical frailty and sarcopenia show increased levels of circulating small extracellular vesicles with a specific mitochondrial signature. Cells 9:973. doi: 10.3390/cells9040973
Picca, A., Calvani, R., Bossola, M., Allocca, E., Menghi, A., Pesce, V., et al. (2018a). Update on mitochondria and muscle aging: all wrong roads lead to sarcopenia. Biol. Chem. 399, 421–436. doi: 10.1515/hsz-2017-0331
Picca, A., Calvani, R., Lorenzi, M., Menghi, A., Galli, M., Vitiello, R., et al. (2017a). Mitochondrial dynamics signaling is shifted toward fusion in muscles of very old hip-fractured patients: results from the Sarcopenia in HIp FracTure (SHIFT) exploratory study. Exp. Gerontol. 96, 63–67. doi: 10.1016/j.exger.2017.06.005
Picca, A., Guerra, F., Calvani, R., Bucci, C., Lo Monaco, M. R., Bentivoglio, A. R., et al. (2019b). Mitochondrial-derived vesicles as candidate biomarkers in Parkinson's disease: rationale, design and methods of the EXosomes in PArkiNson Disease (EXPAND) study. Int. J. Mol. Sci. 20:2373. doi: 10.3390/ijms20102373
Picca, A., Guerra, F., Calvani, R., Marini, F., Biancolillo, A., Landi, G., et al. (2020b). Mitochondrial signatures in circulating extracellular vesicles of older adults with Parkinson's disease: results from the EXosomes in PArkiNson's Disease (EXPAND) study. J. Clin. Med. 9:504. doi: 10.3390/jcm9020504
Picca, A., Lezza, A. M. S., Leeuwenburgh, C., Pesce, V., Calvani, R., Bossola, M., et al. (2018b). Circulating mitochondrial DNA at the crossroads of mitochondrial dysfunction and inflammation during aging and muscle wasting disorders. Rejuvenation Res. 21, 350–359. doi: 10.1089/rej.2017.1989
Picca, A., Lezza, A. M. S., Leeuwenburgh, C., Pesce, V., Calvani, R., Landi, F., et al. (2017b). Fueling inflamm-aging through mitochondrial dysfunction: mechanisms and molecular targets. Int. J. Mol. Sci. 18:933. doi: 10.3390/ijms18050933
Picca, A., Ponziani, F. R., Calvani, R., Marini, F., Biancolillo, A., Coelho-Junior, H. J., et al. (2019a). Gut microbial, inflammatory and metabolic signatures in older people with physical frailty and sarcopenia: results from the BIOSPHERE study. Nutrients 12:65. doi: 10.3390/nu12010065
Ponziani, F. R., Bhoori, S., Castelli, C., Putignani, L., Rivoltini, L., Del Chierico, F., et al. (2019). Hepatocellular carcinoma is associated with gut microbiota profile and inflammation in nonalcoholic fatty liver disease. Hepatology 69, 107–120. doi: 10.1002/hep.30036
Ponziani, F. R., Putignani, L., Paroni Sterbini, F., Petito, V., Picca, A., Del Chierico, F., et al. (2018). Influence of hepatitis C virus eradication with direct-acting antivirals on the gut microbiota in patients with cirrhosis. Aliment Pharmacol. Ther. 48, 1301–1311. doi: 10.1111/apt.15004
Przybyla, B., Gurley, C., Harvey, J. F., Bearden, E., Kortebein, P., Evans, W. J., et al. (2006). Aging alters macrophage properties in human skeletal muscle both at rest and in response to acute resistance exercise. Exp. Gerontol. 41, 320–327. doi: 10.1016/j.exger.2005.12.007
Rockenfeller, P., Koska, M., Pietrocola, F., Minois, N., Knittelfelder, O., Sica, V., et al. (2015). Phosphatidylethanolamine positively regulates autophagy and longevity. Cell Death Differ. 22, 499–508. doi: 10.1038/cdd.2014.219
Roger, J. M., Palagos, B., Bertrand, D., and Fernandez-Ahumada, E. (2011). CovSel: Variable selection for highly multivariate and multi-response calibration. Application to IR spectroscopy. Chemom. Intell. Lab. Syst. 106, 216–223. doi: 10.1016/j.chemolab.2010.10.003
Selathurai, A., Kowalski, G. M., Mason, S. A., Callahan, D. L., Foletta, V. C., Della Gatta, P. A., et al. (2019). Phosphatidylserine decarboxylase is critical for the maintenance of skeletal muscle mitochondrial integrity and muscle mass. Mol. Metab. 27, 33–46. doi: 10.1016/j.molmet.2019.06.020
Sierra, F. (2016). The emergence of geroscience as an interdisciplinary approach to the enhancement of health span and life span. Cold Spring Harb. Perspect. Med. 6:a025163. doi: 10.1101/cshperspect.a025163
Smit, S., van Breemen, M. J., Hoefsloot, H. C. J., Smilde, A. K., Aerts, J. M. F. G., and de Koster, C. G. (2007). Assessing the statistical validity of proteomics based biomarkers. Anal. Chim. Acta 592, 210–217. doi: 10.1016/j.aca.2007.04.043
Soubannier, V., McLelland, G.-L., Zunino, R., Braschi, E., Rippstein, P., Fon, E. A., et al. (2012). A vesicular transport pathway shuttles cargo from mitochondria to lysosomes. Curr. Biol. 22, 135–141. doi: 10.1016/j.cub.2011.11.057
Studenski, S. A., Peters, K. W., Alley, D. E., Cawthon, P. M., McLean, R. R., Harris, T. B., et al. (2014). The FNIH sarcopenia project: rationale, study description, conference recommendations, and final estimates. J. Gerontol. A Biol. Sci. Med. Sci. 69, 547–558. doi: 10.1093/gerona/glu010
Théry, C., Amigorena, S., Raposo, G., and Clayton, A. (2006). Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 3:22. doi: 10.1002/0471143030.cb0322s30
Théry, C., Witwer, K. W., Aikawa, E., Alcaraz, M. J., Anderson, J. D., Andriantsitohaina, R., et al. (2018). Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell Vesicles 7:1535750. doi: 10.1080/20013078.2018.1535750
van der Veen, J. N., Kennelly, J. P., Wan, S., Vance, J. E., Vance, D. E., and Jacobs, R. L. (2017). The critical role of phosphatidylcholine and phosphatidylethanolamine metabolism in health and disease. Biochim. Biophys. Acta Biomembr. 1859, 1558–1572. doi: 10.1016/j.bbamem.2017.04.006
Westerhuis, J. A., Hoefsloot, H. C. J., Smit, S., Vis, D. J., Smilde, A. K., van Velzen, E. J. J., et al. (2008). Assessment of PLSDA cross validation. Metabolomics 4, 81–89. doi: 10.1007/s11306-007-0099-6
Wilson, D., Jackson, T., Sapey, E., and Lord, J. M. (2017). Frailty and sarcopenia: the potential role of an aged immune system. Ageing Res. Reviews 36, 1–10. doi: 10.1016/j.arr.2017.01.006
Yahiaoui, L., Gvozdic, D., Danialou, G., Mack, M., and Petrof, B. J. (2008). CC family chemokines directly regulate myoblast responses to skeletal muscle injury. J. Physiol. 586, 3991–4004. doi: 10.1113/jphysiol.2008.152090
Keywords: aging, biomarkers, cytokines, extracellular vesicles, geroscience, metabolomics, mitochondrial dysfunction, mitochondrial quality control
Citation: Marzetti E, Guerra F, Calvani R, Marini F, Biancolillo A, Gervasoni J, Primiano A, Coelho-Júnior HJ, Landi F, Bernabei R, Bucci C and Picca A (2020) Circulating Mitochondrial-Derived Vesicles, Inflammatory Biomarkers and Amino Acids in Older Adults With Physical Frailty and Sarcopenia: A Preliminary BIOSPHERE Multi-Marker Study Using Sequential and Orthogonalized Covariance Selection – Linear Discriminant Analysis. Front. Cell Dev. Biol. 8:564417. doi: 10.3389/fcell.2020.564417
Received: 21 May 2020; Accepted: 13 August 2020;
Published: 22 September 2020.
Edited by:
Vincenzo Flati, University of L'Aquila, ItalyReviewed by:
Francesco Saverio Dioguardi, University of Milan, ItalyEvasio Pasini, Fondazione Salvatore Maugeri (IRCCS), Italy
Sjors Verlaan, Amsterdam University Medical Center, Netherlands
Copyright © 2020 Marzetti, Guerra, Calvani, Marini, Biancolillo, Gervasoni, Primiano, Coelho-Júnior, Landi, Bernabei, Bucci and Picca. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Riccardo Calvani, cmljY2FyZG8uY2FsdmFuaUBndWVzdC5wb2xpY2xpbmljb2dlbWVsbGkuaXQ=