 Lei-Miao Yin
Lei-Miao Yin Michael Schnoor
Michael Schnoor Chang-Duk Jun
Chang-Duk Jun- 1Laboratory of Molecular Biology, Shanghai Research Institute of Acupuncture and Meridian, Shanghai University of Traditional Chinese Medicine, Shanghai, China
- 2Molecular Biomedicine, Center for Investigation and Advanced Studies of the National Polytechnic Institute (Cinvestav), Mexico City, Mexico
- 3School of Life Sciences, Gwangju Institute of Science and Technology, Gwangju, South Korea
The calponin homology (CH) domain is one of the most common modules in various actin-binding proteins and is characterized by an α-helical fold. The CH domain plays important regulatory roles in both cytoskeletal dynamics and signaling. The CH domain is required for stability and organization of the actin cytoskeleton, calcium mobilization and activation of downstream pathways. The CH domain has recently garnered increased attention due to its importance in the onset of different diseases, such as cancers and asthma. However, many roles of the CH domain in various protein functions and corresponding diseases are still unclear. Here, we review current knowledge about the structural features, interactome and related diseases of the CH domain.
Introduction
Actin is an essential cytoskeletal protein that plays a critical role in multiple cellular processes (Pollard and Goldman, 2018). Actin monomers are assembled into different filamentous structures to form the actin cytoskeleton, which is a highly dynamic structure that regulates many cell processes such as adhesion, spreading and migration. Actin cytoskeletal dynamics require the coordinated action of many different actin-binding proteins (ABPs) (Garcia-Ponce et al., 2015). Since the discovery of actin-binding proteins, such as α-actinin and filamin, filamin in the 1970s (Lazarides and Burridge, 1975; Shizuta et al., 1976), more than 160 different members have been identified (Lappalainen, 2016; Kuhn and Mannherz, 2017). Calponin is an ABP that is expressed in smooth muscle and multiple types of non-muscle cells (Liu and Jin, 2016). The calponin homology (CH) domain, first identified at the N-terminus of calponin, is a common peptide module of approximately 100 residues and its precise number varies from protein to protein (Castresana and Saraste, 1995; Korenbaum and Rivero, 2002; Gimona and Winder, 2008). Sequence alignment of the CH domain shows that the residues of tryptophan (W) in helix I and aspartate (D) in helix VI are the most-conserved residues, while the consensus motif DGXXLXXL appears in helix III (Figure 1A; Gimona and Winder, 2008). The CH domain has been identified in a variety of proteins (CH-domain-containing proteins, CCPs), whose functions range from actin cross-linking to signal regulation (Gimona and Winder, 2008). Three types of CH domains have been described mainly based on their functions (Gimona et al., 2002; Korenbaum and Rivero, 2002). The type 1 CH domain (CH1) has the intrinsic ability to bind to F-actin. The type 2 CH (CH2) domain binds in tandem with CH1 and is required to facilitate high-affinity binding of F-actin. The type 3 CH domain (CH3) usually acts as a single CH domain in several ABPs and signaling proteins. Despite similarity of the secondary structure with the other types of CH domains, the CH3 domain shows functional diversity due to its ability to interact with many different proteins (Stradal et al., 1998).
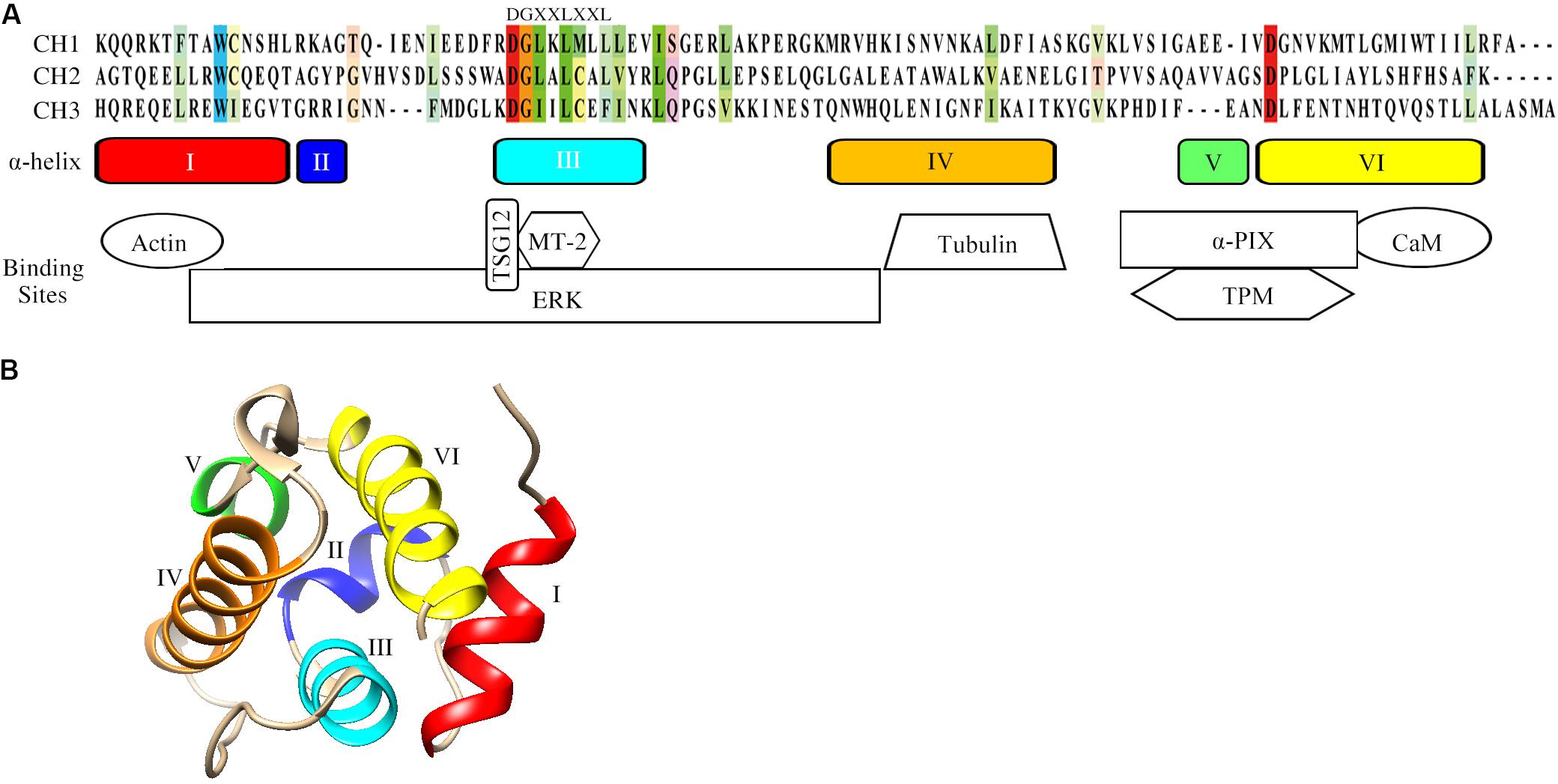
Figure 1. Structural characteristics of the CH domain. (A) Sequence alignment, schematic of the secondary structure elements and the binding sites of CH domain for actin and signaling proteins. The conserved residues among the three CH domains are colored. Schematics of the secondary structure elements of CH domain and binding sites are also included. UniProt identifiers for CH1 (α-actinin), CH2 (MICAL, Molecule interacting with CasL) and CH3 (calponin-1) are P12814, Q8TDZ2, P51911. (B) The tertiary fold of the calponin CH domain (PDB: 1H67). The CH domain contains in total six α-helices. Helices III and VI are approximately parallel, while helix IV is lying oblique aside. The structural model was generated by UCSF Chimera. Abbreviations: αPIX: Cdc42/Rac1-specific guanine nucleotide exchanging factor; CaM: calmodulin; ERK: extracellular signal-regulated kinase; MT-2: metallothionein-2; TPM: tropomyosin; TSG12: a specific transgelin-2 agonist.
Recent studies have reported that CCPs, including molecule interacting with CasL (MICAL), leucine-rich repeats and calponin homology containing 4 (Lrch4), smoothelin-like 1 (SMTNL1) and transgelin-2, exhibit an unappreciated functional variety and play important roles in the onset of various diseases (Table 1; Yin et al., 2018; Min et al., 2019). However, the precise role of how the CH domain confers various protein functions to drive such diseases is still unclear. In this review, we summarize the structural features, binding partners and diseases related to the CH domain.
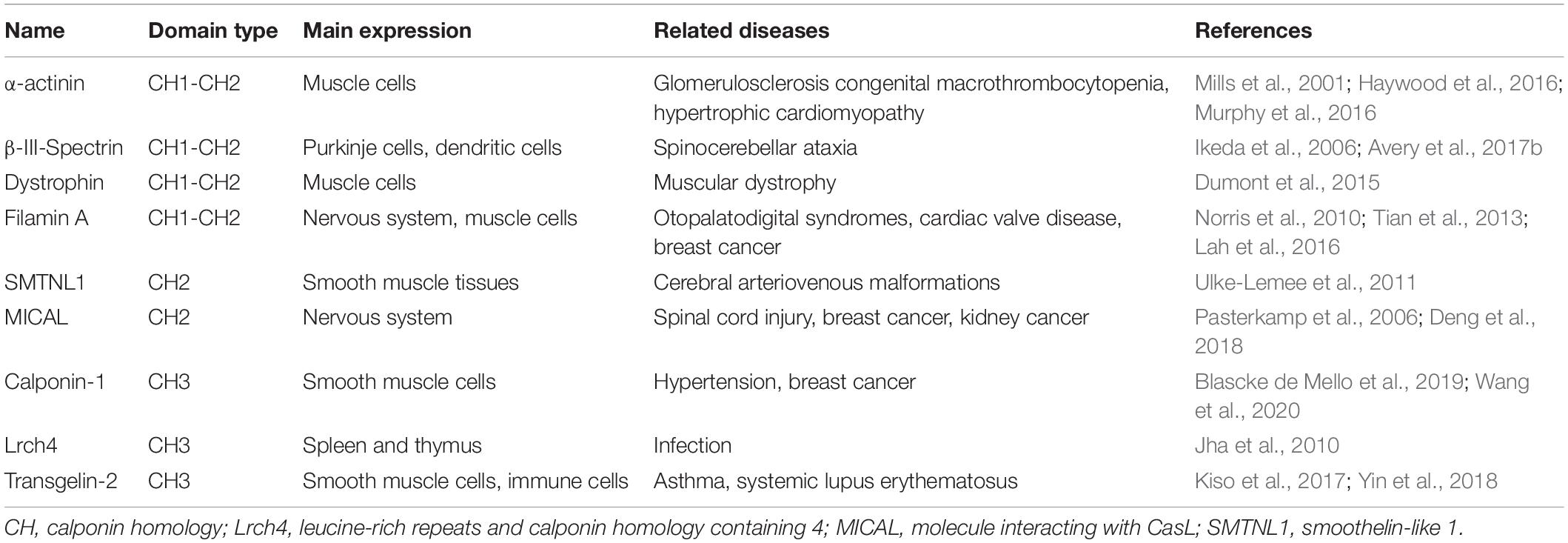
Table 1. CH-domain-containing proteins involved in the onset and progression of different diseases.
The Structure of the CH Domain
Understanding common CH domain structural features can help to elucidate the functions of CCPs. The CH domain is mainly α-helical and the strictly conserved residues in α-helices constitute an invariant hydrophobic core (Korenbaum and Rivero, 2002). The tertiary structure of the CH domain is compact and maintained by a network of hydrophobic interactions (Bramham et al., 2002). The CH domain contains six α-helices in total, including a core of four α-helices (I, III, IV, and VI) and two short helical structures (II and V, Figure 1A). A 310-helical turn is also present in the loop between helices IV and V. Three helices of the CH domain of calponins (III, IV, and VI) form a triple-helix bundle, and helix I binds at a right angle across the surface provided by helices III and VI (Figure 1B). Unlike the majority of the CH domains, which are generally located at the N-terminus of proteins, the single CH2 domain of SMTNL1 is at its C-terminus. Nuclear magnetic resonance data show that the CH2 domain of SMTNL1 adopts the same α-helical fold as other CH domains and the most notable difference is the “KTKKK” cluster in the final helix (Ishida et al., 2008). The cluster “KTKKK” is unique in SMTNL1 and may be the potential site for ubiquitinylation (Ishida et al., 2008).
Functional Regulation of the CH Domain
Changes in the linker and flanking regions of the CH domain regulate the configuration of the domains, thus influencing functional regulation and affinity for their interaction partners. A hinge (GLQQTN) in the linker region between the CH1-CH2 domain of dystrophin acts like a swivel to allow these conformational transitions (Fealey et al., 2018). After the linkers of the dystrophin and utrophin tandem CH domains were swapped, the dystrophin tandem CH domain with an utrophin linker (DUL) showed a 2-fold higher binding affinity compared to that of the dystrophin tandem CH domain, while the utrophin tandem CH domain with a dystrophin linker (UDL) had a 50% lower binding affinity than the utrophin tandem CH domain (Bandi et al., 2015). A chimera containing the CH1-CH2 domain from utrophin and the linker region from filamin A had a significantly higher actin-binding affinity (KD = 0.7 μM) than wild-type utrophin (KD = 19 μM) (Harris et al., 2019). Moreover, the N-terminal flanking region of the CH domain influences the binding of F-actin. After truncation of a flanking region of utrophin (MAKYGEHEASPDNGQNEFSDIIKSRSD), the binding affinity for F-actin was significantly decreased by approximately 2-fold in HeLa and HEK293T cells (Harris et al., 2019). However, in vitro co-sedimentation assays (not in live cells) showed that the same truncated utrophin binds to F-actin 30 times weaker than the full-length protein (Singh et al., 2017).
Binding Partners of the CH Domain
The functional diversity of different CH domains is a result of different binding partners including actin, tubulin and signaling proteins as described below (Galkin et al., 2006).
Binding With Actin and Tubulin
The exact mechanisms regulating binding between actin and the CH domain are still unclear. This process is associated with complex factors, including the CH domain number (single or a tandem pair), conformational differences, and flanking sequences (Galkin et al., 2010; Harris et al., 2019). The CH domain mainly acts in tandem pairs for F-actin binding (Sjoblom et al., 2008). Cryo-electron microscopy data revealed that the CH1 domain of human filamin A contributed to F-actin binding without direct CH2 and actin interactions (Iwamoto et al., 2018). Binding of the CH1-CH2 domain and actin is mechanically regulated via closed or open conformations (Shams et al., 2016). The CH1 domain contains the main actin-binding sites, however, one of the binding sites of CH1 is buried within the CH1-CH2 interface and only becomes accessible in the open conformation (Borrego-Diaz et al., 2006). Therefore, the CH2 domain serves as a locator domain to position the true actin-binding motifs, including the regulation of CH1 binding with actin, prevention of actin clashes and stabilization of the actin-binding domain (Galkin et al., 2006; Iwamoto et al., 2018). Co-sedimentation assays showed that the binding to F-actin by a single CH1 domain of human utrophin was about 5-fold weaker than that of the CH tandem pair, while both binding constants were 1000-fold stronger than that of the single CH2 domain (Singh et al., 2014).
Residue mutations can also affect binding between the CH domain and F-actin. For example, the CH2 domain mutation (L253P) of β-III-spectrin caused opening of the CH1-CH2 domains and promoted the N-terminal region of CH1 to become α-helical, thus enhancing approximately 1000-fold the actin-binding affinity (Avery et al., 2017a). In contrast, a single cysteine mutation (C10S before the CH1 domain or C188S in the middle of the CH2 domain) did not affect the structure or stability of the CH1-CH2 domain of dystrophin (Singh and Mallela, 2012). However, the K237E mutation in CH2 of α-actinin decreased the open conformation strength of the CH domain and increased actin-binding affinity (Shams et al., 2016).
Whether a single CH domain binds actin is still controversial. By constructing calponin without C-terminal tandem repeats, the resulting protein with the CH domain failed to bind to actin (Gimona and Mital, 1998), suggesting that a single CH domain is neither sufficient nor necessary for the binding of F-actin (Gimona and Mital, 1998). However, new findings showed opposite results within cells. Transgelin-1 interacted with actin via its CH3 domain, while the C-terminal tandem repeats were dispensable for actin-binding in smooth muscle cells (Matsui et al., 2018). While the wild-type transgelin-1 or transgelin-1 without the C-terminal tandem repeats both displayed fibrous patterns, the truncated protein with deletion of the CH3 domain showed diffuse patterns after separate transfection into A7r5 smooth muscle cells (Matsui et al., 2018).
The CH domain can also bind other cytoskeletal proteins such as tubulin. The end-binding protein 1 (EB1) is the first example of a single CH domain that can associate with tubulin (Hayashi and Ikura, 2003). A truncated version of EB1 containing only the CH domain co-sedimented with tubulin (Hayashi and Ikura, 2003). In contrast, the mutation K89E within α-helix IV, close to the hydrophobic cleft of the CH domain, abolished tubulin binding (Hayashi and Ikura, 2003). Deletion of the N-terminal 207 amino acid region of Hec1 (i.e., Hec1 without CH domain) resulted in failure of chromosomes alignment at the spindle equator during mitosis in PtK1 cells (Guimaraes et al., 2008). By contrast, deletion of only the N-terminal 80 amino acid tail of Hec1 (i.e., Hec1 with the CH domain) did not affect protein function. These findings together suggested that the CH domain of Hec1 is required for efficient binding of tubulin (Guimaraes et al., 2008). These new evidences clearly show the distinctive binding mechanisms of the CH domains with actin and tubulin, highlighting the need for further investigation into the functional mechanisms of these binding patterns.
Binding With Signaling Proteins
Besides its ability to bind to actin and tubulin, the CH domain can participate in signal transduction by binding to different protein partners such as extracellular regulated kinase (ERK) and calmodulin (Figure 1A). The CH domain of calponin was identified as the binding site for ERK by sequencing chymotryptic fragments of calponin (Leinweber et al., 1999). Calponin thus facilitates the formation of signaling complexes with ERK and other kinases, such as protein kinase C (Leinweber et al., 2000). SMTNL1 can also interact with signaling proteins, including calmodulin and tropomyosin. The sequence IQELYRSLVQK in the α-helix VI of the SMTNL1 CH2 domain is the binding site for calmodulin and the KD obtained by isothermal titration calorimetry was 2.7 × 10–6 M (Ishida et al., 2008). SMTNL1 can be phosphorylated by protein kinase A (PKA) at Ser301, which lies upstream of the CH domain, and this phosphorylation strongly enhances the ability of SMTNL1 to associate with tropomyosin (Ulke-Lemee et al., 2017). However, the exact binding region between SMTNL1 and tropomyosin that may affect the modulation of muscle contractile activity is still uncertain (Ulke-Lemee et al., 2010). Removal of the CH2 domain or expression of the CH2 domain of SMTNL1 alone did not enable binding with tropomyosin, suggesting that the CH2 domain is not sufficient to mediate binding but is involved in the regulation of the binding affinity for tropomyosin (Ulke-Lemee et al., 2010). However, the study further shows that a portion of the N-terminal intrinsically disordered region (1–341 residues) of SMTNL1 forms intramolecular contacts with its C-terminal CH domain, SMTNL1 thus interacts with tropomyosin at residues 421–436, which encompasses the entirety of α-helix V and the beginning of α-helix VI of the CH2 domain (MacDonald et al., 2012).
For other CCPs, the N-terminal 53–271 residues of affixin that cover the CH1 domain but not the CH2 domain are the binding sites of Cdc42/Rac1-specific guanine nucleotide exchanging factor (αPIX), as shown using co-immunoprecipitation assays (Mishima et al., 2004). Transgelin-2 with a CH3 domain is a receptor for extracellular ligands such as metallothionein-2 (Crunkhorn, 2018; Yin et al., 2019). The small compound TSG12, which was identified through molecular docking by targeting 46–63 residues of the CH3 domain of human transgelin-2 (QPGRENFQKWLKDGTVLC) induced dephosphorylation of myosin phosphatase-targeting subunit 1 (MYPT1) (Yin et al., 2018).
The Role of CH Domains in Various Diseases
CCPs, including MICAL1/2, Lrch4 and SMTNL1, have been shown to play crucial roles in various diseases as discussed below. A summary of the involvement of CH domains in various diseases is shown in Table 1.
MICAL1/2
MICAL1/2 contains a CH2 domain and oxidizes methionine residues of actin to disassemble F-actin into G-actin (Grintsevich et al., 2016). The MICAL1/2 CH2 domain is connected to the monooxygenase domain, and Arg530 in the CH2 domain is the key residue mediating interaction with the monooxygenase domain (Kim et al., 2020). MICAL1/2 can also regulate actin dynamics and cell morphological changes via the CH2 domain through interacting with other signaling proteins (Hung et al., 2010).
Studies have shown that MICAL proteins are closely related to neural diseases and cancers. MICAL expression is substantially elevated in oligodendrocytes and in meningeal fibroblasts during spinal cord injury, suggesting an involvement of MICAL in neuronal regeneration (Pasterkamp et al., 2006). Targeting MICAL may provide a new therapeutic option for cancer treatment (Yoon and Terman, 2018). For example, deletion of MICAL1 substantially reduced cell proliferation in the breast cancer cell lines MCF-7 and T47D (Deng et al., 2018). Over-expression of MICAL2 in MCF-7 cells augmented the level of epidermal growth factor receptor (EGFR) in the plasma membrane, thus enhancing cell migration (Wang et al., 2018). In contrast, silencing MICAL2 in MDA-MB-231 cells degraded EGFR and inhibited cell migration (Wang et al., 2018). MICAL2 gene expression was significantly increased in aggressive primary gastric and renal cancers (Mariotti et al., 2016). MICAL2 knockdown caused a reduction in viability and loss of motility and invasion in 786-O kidney cancer cells, suggesting that MICAL2 might be a promising target for anti-metastatic therapy (Mariotti et al., 2016).
Lrch4
Lrch4 is a plasma membrane protein abundantly expressed in the spleen and thymus, containing a single-pass transmembrane domain with nine leucine-rich repeats and a CH3 domain in its ectodomain (Aloor et al., 2019). Recent data showed that Lrch4 did not interact with ezrin, radixin and moesin (ERM) in drosophila S2 cells, suggesting that the CH3 domain of Lrch4 may not bind with the FERM domain of ERM (Foussard et al., 2010). The function of the CH3 domain in Lrch4 is still unclear (Aloor et al., 2019).
Lrch4 is a novel Toll-like receptor (TLR) accessory protein as Lrch4 knockdown attenuated TNFα secretion induced by various TLR ligands (Aloor et al., 2019). Therefore, Lrch4 has been considered a broad-spanning regulator of the innate immune response and a potential molecular target in inflammatory diseases (Aloor et al., 2019). Lrch4 was identified by mass spectrometry to be differentially expressed in macrophages 24 h after infection with Mycobacterium avium subsp hominissuis (Jha et al., 2010). Microarray analysis showed that the gene expression of Lrch4 was up-regulated by 1.6-fold in lipopolysaccharide-stimulated dendritic cells in inflammation (Ceppi et al., 2009).
SMTNL1
Smoothelin-like 1, which contains a CH2 domain in the C-terminal region, is a novel member of the smoothelin protein family (Borman et al., 2004). Deletion of the CH2 domain can significantly change the intracellular localization of SMTNL1 (from distributed longitudinally on F-actin to diffuse distribution in the cytoplasm) in rat vascular smooth muscle cells suggesting that the CH2 domain is critical for F-actin binding (Ulke-Lemee et al., 2010). Experiments with truncated recombinant proteins showed that the CH2 domain was essential for SMTNL1-associated smooth muscle relaxation because the CH2 domain alone did not cause relaxation in rabbit ileum smooth muscle strips (Borman et al., 2009).
Smoothelin-like 1 can modulate muscle contractility, and its biological activity may involve interaction with the contractile actin machinery (Ulke-Lemee et al., 2010). One of the target genes of SMTNL1 is MYPT1 (high expression of MYPT1 is associated with the contraction of smooth muscle), and SMTNL1 knock-out increased MYPT1 protein expression by 30- to 40-fold in neonatal mice (Lontay et al., 2010). SMTNL1 interacts with myosin phosphatase in the cytoplasm, however, when phosphorylated at Ser301 in response to PKA/PKC, SMTNL1 translocates into the nucleus where it may activate transcription factors driving MYPT1 expression (Lontay et al., 2010). The gene expression of SMTNL1 was also significantly increased by approximately 4-fold in human cerebral arteriovenous malformations, suggesting that the elevated level of SMTNL1 may decrease MYPT1 expression to relax brain blood vessels and thus contribute to this lumen disorder (Yao et al., 2019).
Conclusion and Prospects
The CH domain displays high structural conservation, but shows diverse biological functions. The indispensable flanking regions and/or intrinsically unfolded protein modules may contribute to orchestrating CH domain functions. However, when comparing cell-based experiments with in vitro experiments using recombinant proteins only, it should be noted that due to the complexities of the cellular environment, other factors could be coming into play and distorting the results. With newly discovered proteins that interact with CH domains, some of the diverse functions have now been elucidated. However, many other binding proteins and functions certainly remain to be discovered, thus warranting further research into CH domain biology. Given that numerous CCPs, such as MICAL and transgelin-2, have been identified as promising therapeutic targets in diseases, it will be important to investigate in the future whether compounds can be designed to specifically target CH domains and thus improve the outcome of certain diseases.
Author Contributions
L-MY designed the work, wrote the manuscript, and prepared the figures. MS and C-DJ drafted and revised the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
Funding
This work was supported by National Natural Science Foundation of China (Nos. 81922076, 81873373, and 81973951).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Aloor, J. J., Azzam, K. M., Guardiola, J. J., Gowdy, K. M., Madenspacher, J. H., Gabor, K. A., et al. (2019). Leucine-rich repeats and calponin homology containing 4 (Lrch4) regulates the innate immune response. J. Biol. Chem. 294, 1997–2008. doi: 10.1074/jbc.RA118.004300
Avery, A. W., Fealey, M. E., Wang, F., Orlova, A., Thompson, A. R., Thomas, D. D., et al. (2017a). Structural basis for high-affinity actin binding revealed by a beta-III-spectrin SCA5 missense mutation. Nat. Commun. 8:1350. doi: 10.1038/s41467-017-01367-w
Avery, A. W., Thomas, D. D., and Hays, T. S. (2017b). beta-III-spectrin spinocerebellar ataxia type 5 mutation reveals a dominant cytoskeletal mechanism that underlies dendritic arborization. Proc. Natl. Acad. Sci. U.S.A. 114, E9376–E9385. doi: 10.1073/pnas.1707108114
Bandi, S., Singh, S. M., and Mallela, K. M. (2015). Interdomain linker determines primarily the structural stability of dystrophin and utrophin tandem calponin-homology domains rather than their actin-binding affinity. Biochemistry 54, 5480–5488. doi: 10.1021/acs.biochem.5b00741
Blascke de Mello, M. M., Parente, J. M., Schulz, R., and Castro, M. M. (2019). Matrix metalloproteinase (MMP)-2 activation by oxidative stress decreases aortic calponin-1 levels during hypertrophic remodeling in early hypertension. Vascul Pharmacol. 116, 36–44. doi: 10.1016/j.vph.2018.10.002
Borman, M. A., Freed, T. A., Haystead, T. A., and Macdonald, J. A. (2009). The role of the calponin homology domain of smoothelin-like 1 (SMTNL1) in myosin phosphatase inhibition and smooth muscle contraction. Mol. Cell Biochem. 327, 93–100. doi: 10.1007/s11010-009-0047-z
Borman, M. A., MacDonald, J. A., and Haystead, T. A. (2004). Modulation of smooth muscle contractility by CHASM, a novel member of the smoothelin family of proteins. FEBS Lett. 573, 207–213. doi: 10.1016/j.febslet.2004.08.002
Borrego-Diaz, E., Kerff, F., Lee, S. H., Ferron, F., Li, Y., and Dominguez, R. (2006). Crystal structure of the actin-binding domain of alpha-actinin 1: evaluating two competing actin-binding models. J. Struct. Biol. 155, 230–238. doi: 10.1016/j.jsb.2006.01.013
Bramham, J., Hodgkinson, J. L., Smith, B. O., Uhrin, D., Barlow, P. N., and Winder, S. J. (2002). Solution structure of the calponin CH domain and fitting to the 3D-helical reconstruction of F-actin:calponin. Structure 10, 249–258. doi: 10.1016/s0969-2126(02)00703-7
Castresana, J., and Saraste, M. (1995). Does vav bind to f-actin through a CH domain? FEBS Lett. 374, 149–151. doi: 10.1016/0014-5793(95)01098-y
Ceppi, M., Pereira, P. M., Dunand-Sauthier, I., Barras, E., Reith, W., Santos, M. A., et al. (2009). MicroRNA-155 modulates the interleukin-1 signaling pathway in activated human monocyte-derived dendritic cells. Proc. Natl. Acad. Sci. U.S.A. 106, 2735–2740. doi: 10.1073/pnas.0811073106
Crunkhorn, S. (2018). Asthma: alternative route to airway relaxation. Nat. Rev. Drug Discov. 17:241. doi: 10.1038/nrd.2018.37
Deng, W., Wang, Y., Zhao, S., Zhang, Y., Chen, Y., Zhao, X., et al. (2018). MICAL1 facilitates breast cancer cell proliferation via ROS-sensitive ERK/cyclin D pathway. J. Cell Mol. Med. 22, 3108–3118. doi: 10.1111/jcmm.13588
Dumont, N. A., Wang, Y. X., von Maltzahn, J., Pasut, A., Bentzinger, C. F., Brun, C. E., et al. (2015). Dystrophin expression in muscle stem cells regulates their polarity and asymmetric division. Nat. Med. 21, 1455–1463. doi: 10.1038/nm.3990
Fealey, M. E., Horn, B., Coffman, C., Miller, R., Lin, A. Y., Thompson, A. R., et al. (2018). Dynamics of dystrophin’s actin-binding domain. Biophys. J. 115, 445–454. doi: 10.1016/j.bpj.2018.05.039
Foussard, H., Ferrer, P., Valenti, P., Polesello, C., Carreno, S., and Payre, F. (2010). LRCH proteins: a novel family of cytoskeletal regulators. PLoS ONE 5:e12257. doi: 10.1371/journal.pone.0012257
Galkin, V. E., Orlova, A., Fattoum, A., Walsh, M. P., and Egelman, E. H. (2006). The CH-domain of calponin does not determine the modes of calponin binding to F-actin. J. Mol. Biol. 359, 478–485. doi: 10.1016/j.jmb.2006.03.044
Galkin, V. E., Orlova, A., Salmazo, A., Djinovic-Carugo, K., and Egelman, E. H. (2010). Opening of tandem calponin homology domains regulates their affinity for F-actin. Nat. Struct. Mol. Biol. 17, 614–616. doi: 10.1038/nsmb.1789
Garcia-Ponce, A., Citalan-Madrid, A. F., Velazquez-Avila, M., Vargas-Robles, H., and Schnoor, M. (2015). The role of actin-binding proteins in the control of endothelial barrier integrity. Thromb Haemost. 113, 20–36. doi: 10.1160/TH14-04-0298
Gimona, M., Djinovic-Carugo, K., Kranewitter, W. J., and Winder, S. J. (2002). Functional plasticity of CH domains. FEBS Lett. 513, 98–106. doi: 10.1016/s0014-5793(01)03240-9
Gimona, M., and Mital, R. (1998). The single CH domain of calponin is neither sufficient nor necessary for F-actin binding. J. Cell Sci. 111(Pt 13), 1813–1821.
Gimona, M., and Winder, S. J. (2008). “The calponin homology (CH) domain,” in Protein Science Encyclopedia, ed. A. Fersht (Weinham: Wiley-VCH Verlag GmbH & Co. KGaA), 1–16. doi: 10.1002/9783527610754
Grintsevich, E. E., Yesilyurt, H. G., Rich, S. K., Hung, R. J., Terman, J. R., and Reisler, E. (2016). F-actin dismantling through a redox-driven synergy between Mical and cofilin. Nat. Cell Biol. 18, 876–885. doi: 10.1038/ncb3390
Guimaraes, G. J., Dong, Y., McEwen, B. F., and Deluca, J. G. (2008). Kinetochore-microtubule attachment relies on the disordered N-terminal tail domain of Hec1. Curr. Biol. 18, 1778–1784. doi: 10.1016/j.cub.2008.08.012
Harris, A. R., Belardi, B., Jreij, P., Wei, K., Shams, H., Bausch, A., et al. (2019). Steric regulation of tandem calponin homology domain actin-binding affinity. Mol. Biol. Cell 30, 3112–3122. doi: 10.1091/mbc.E19-06-0317
Hayashi, I., and Ikura, M. (2003). Crystal structure of the amino-terminal microtubule-binding domain of end-binding protein 1 (EB1). J. Biol. Chem. 278, 36430–36434. doi: 10.1074/jbc.M305773200
Haywood, N. J., Wolny, M., Rogers, B., Trinh, C. H., Shuping, Y., Edwards, T. A., et al. (2016). Hypertrophic cardiomyopathy mutations in the calponin-homology domain of ACTN2 affect actin binding and cardiomyocyte Z-disc incorporation. Biochem. J. 473, 2485–2493. doi: 10.1042/BCJ20160421
Hung, R. J., Yazdani, U., Yoon, J., Wu, H., Yang, T., Gupta, N., et al. (2010). Mical links semaphorins to F-actin disassembly. Nature 463, 823–827. doi: 10.1038/nature08724
Ikeda, Y., Dick, K. A., Weatherspoon, M. R., Gincel, D., Armbrust, K. R., Dalton, J. C., et al. (2006). Spectrin mutations cause spinocerebellar ataxia type 5. Nat. Genet. 38, 184–190. doi: 10.1038/ng1728
Ishida, H., Borman, M. A., Ostrander, J., Vogel, H. J., and MacDonald, J. A. (2008). Solution structure of the calponin homology (CH) domain from the smoothelin-like 1 protein: a unique apocalmodulin-binding mode and the possible role of the C-terminal type-2 CH-domain in smooth muscle relaxation. J. Biol. Chem. 283, 20569–20578. doi: 10.1074/jbc.M800627200
Iwamoto, D. V., Huehn, A., Simon, B., Huet-Calderwood, C., Baldassarre, M., Sindelar, C. V., et al. (2018). Structural basis of the filamin A actin-binding domain interaction with F-actin. Nat. Struct. Mol. Biol. 25, 918–927. doi: 10.1038/s41594-018-0128-3
Jha, S. S., Danelishvili, L., Wagner, D., Maser, J., Li, Y. J., Moric, I., et al. (2010). Virulence-related Mycobacterium avium subsp hominissuis MAV_2928 gene is associated with vacuole remodeling in macrophages. BMC Microbiol. 10:100. doi: 10.1186/1471-2180-10-100
Kim, J., Lee, H., Roh, Y. J., Kim, H. U., Shin, D., Kim, S., et al. (2020). Structural and kinetic insights into flavin-containing monooxygenase and calponin-homology domains in human MICAL3. IUCrJ 7(Pt 1), 90–99. doi: 10.1107/S2052252519015409
Kiso, K., Yoshifuji, H., Oku, T., Hikida, M., Kitagori, K., Hirayama, Y., et al. (2017). Transgelin-2 is upregulated on activated B-cells and expressed in hyperplastic follicles in lupus erythematosus patients. PLoS ONE 12:e0184738. doi: 10.1371/journal.pone.0184738
Korenbaum, E., and Rivero, F. (2002). Calponin homology domains at a glance. J. Cell Sci. 115(Pt 18), 3543–3545. doi: 10.1242/jcs.00003
Kuhn, S., and Mannherz, H. G. (2017). Actin: structure, function, dynamics, and interactions with bacterial toxins. Curr. Top. Microbiol. Immunol. 399, 1–34. doi: 10.1007/82_2016_45
Lah, M., Niranjan, T., Srikanth, S., Holloway, L., Schwartz, C. E., Wang, T., et al. (2016). A distinct X-linked syndrome involving joint contractures, keloids, large optic cup-to-disc ratio, and renal stones results from a filamin A (FLNA) mutation. Am. J. Med. Genet. A 170A, 881–890. doi: 10.1002/ajmg.a.37567
Lappalainen, P. (2016). Actin-binding proteins: the long road to understanding the dynamic landscape of cellular actin networks. Mol. Biol. Cell 27, 2519–2522. doi: 10.1091/mbc.E15-10-0728
Lazarides, E., and Burridge, K. (1975). Alpha-actinin: immunofluorescent localization of a muscle structural protein in nonmuscle cells. Cell 6, 289–298. doi: 10.1016/0092-8674(75)90180-4
Leinweber, B., Parissenti, A. M., Gallant, C., Gangopadhyay, S. S., Kirwan-Rhude, A., Leavis, P. C., et al. (2000). Regulation of protein kinase C by the cytoskeletal protein calponin. J. Biol. Chem. 275, 40329–40336. doi: 10.1074/jbc.M008257200
Leinweber, B. D., Leavis, P. C., Grabarek, Z., Wang, C. L., and Morgan, K. G. (1999). Extracellular regulated kinase (ERK) interaction with actin and the calponin homology (CH) domain of actin-binding proteins. Biochem. J. 344(Pt 1), 117–123.
Liu, R., and Jin, J. P. (2016). Calponin isoforms CNN1, CNN2 and CNN3: regulators for actin cytoskeleton functions in smooth muscle and non-muscle cells. Gene 585, 143–153. doi: 10.1016/j.gene.2016.02.040
Lontay, B., Bodoor, K., Weitzel, D. H., Loiselle, D., Fortner, C., Lengyel, S., et al. (2010). Smoothelin-like 1 protein regulates myosin phosphatase-targeting subunit 1 expression during sexual development and pregnancy. J. Biol. Chem. 285, 29357–29366. doi: 10.1074/jbc.M110.143966
MacDonald, J. A., Ishida, H., Butler, E. I., Ulke-Lemee, A., Chappellaz, M., Tulk, S. E., et al. (2012). Intrinsically disordered N-terminus of calponin homology-associated smooth muscle protein (CHASM) interacts with the calponin homology domain to enable tropomyosin binding. Biochemistry 51, 2694–2705. doi: 10.1021/bi2019018
Mariotti, S., Barravecchia, I., Vindigni, C., Pucci, A., Balsamo, M., Libro, R., et al. (2016). MICAL2 is a novel human cancer gene controlling mesenchymal to epithelial transition involved in cancer growth and invasion. Oncotarget 7, 1808–1825. doi: 10.18632/oncotarget.6577
Matsui, T. S., Ishikawa, A., and Deguchi, S. (2018). Transgelin-1 (SM22alpha) interacts with actin stress fibers and podosomes in smooth muscle cells without using its actin binding site. Biochem. Biophys. Res. Commun. 505, 879–884. doi: 10.1016/j.bbrc.2018.09.176
Mills, M., Yang, N., Weinberger, R., Vander Woude, D. L., Beggs, A. H., Easteal, S., et al. (2001). Differential expression of the actin-binding proteins, alpha-actinin-2 and -3, in different species: implications for the evolution of functional redundancy. Hum. Mol. Genet. 10, 1335–1346. doi: 10.1093/hmg/10.13.1335
Min, P., Zhao, S., Liu, L., Zhang, Y., Ma, Y., Zhao, X., et al. (2019). MICAL-L2 potentiates Cdc42-dependent EGFR stability and promotes gastric cancer cell migration. J. Cell. Mol. Med. 23, 4475–4488. doi: 10.1111/jcmm.14353
Mishima, W., Suzuki, A., Yamaji, S., Yoshimi, R., Ueda, A., Kaneko, T., et al. (2004). The first CH domain of affixin activates Cdc42 and Rac1 through alphaPIX, a Cdc42/Rac1-specific guanine nucleotide exchanging factor. Genes Cells 9, 193–204. doi: 10.1111/j.1356-9597.2004.00717.x
Murphy, A. C., Lindsay, A. J., McCaffrey, M. W., Djinovic-Carugo, K., and Young, P. W. (2016). Congenital macrothrombocytopenia-linked mutations in the actin-binding domain of alpha-actinin-1 enhance F-actin association. FEBS Lett. 590, 685–695. doi: 10.1002/1873-3468.12101
Norris, R. A., Moreno-Rodriguez, R., Wessels, A., Merot, J., Bruneval, P., Chester, A. H., et al. (2010). Expression of the familial cardiac valvular dystrophy gene, filamin-A, during heart morphogenesis. Dev. Dyn. 239, 2118–2127. doi: 10.1002/dvdy.22346
Pasterkamp, R. J., Dai, H. N., Terman, J. R., Wahlin, K. J., Kim, B., Bregman, B. S., et al. (2006). MICAL flavoprotein monooxygenases: expression during neural development and following spinal cord injuries in the rat. Mol. Cell. Neurosci. 31, 52–69. doi: 10.1016/j.mcn.2005.09.001
Pollard, T. D., and Goldman, R. D. (2018). Overview of the cytoskeleton from an evolutionary perspective. Cold Spring. Harb. Perspect. Biol. 10:a030288. doi: 10.1101/cshperspect.a030288
Shams, H., Golji, J., Garakani, K., and Mofrad, M. R. (2016). Dynamic regulation of alpha-actinin’s calponin homology domains on F-actin. Biophys. J. 110, 1444–1455. doi: 10.1016/j.bpj.2016.02.024
Shizuta, Y., Shizuta, H., Gallo, M., Davies, P., and Pastan, I. (1976). Purification and properties of filamin, and actin binding protein from chicken gizzard. J. Biol. Chem. 251, 6562–6567.
Singh, S. M., Bandi, S., and Mallela, K. M. (2017). The N-terminal flanking region modulates the actin binding affinity of the utrophin tandem calponin-homology domain. Biochemistry 56, 2627–2636. doi: 10.1021/acs.biochem.6b01117
Singh, S. M., Bandi, S., Winder, S. J., and Mallela, K. M. (2014). The actin binding affinity of the utrophin tandem calponin-homology domain is primarily determined by its N-terminal domain. Biochemistry 53, 1801–1809. doi: 10.1021/bi500149q
Singh, S. M., and Mallela, K. M. (2012). The N-terminal actin-binding tandem calponin-homology (CH) domain of dystrophin is in a closed conformation in solution and when bound to F-actin. Biophys. J. 103, 1970–1978. doi: 10.1016/j.bpj.2012.08.066
Sjoblom, B., Ylanne, J., and Djinovic-Carugo, K. (2008). Novel structural insights into F-actin-binding and novel functions of calponin homology domains. Curr. Opin. Struct. Biol. 18, 702–708. doi: 10.1016/j.sbi.2008.10.003
Stradal, T., Kranewitter, W., Winder, S. J., and Gimona, M. (1998). CH domains revisited. FEBS Lett. 431, 134–137. doi: 10.1016/s0014-5793(98)00751-0
Tian, H. M., Liu, X. H., Han, W., Zhao, L. L., Yuan, B., and Yuan, C. J. (2013). Differential expression of filamin A and its clinical significance in breast cancer. Oncol. Lett. 6, 681–686. doi: 10.3892/ol.2013.1454
Ulke-Lemee, A., Ishida, H., Borman, M. A., Valderrama, A., Vogel, H. J., and MacDonald, J. A. (2010). Tropomyosin-binding properties of the CHASM protein are dependent upon its calponin homology domain. FEBS Lett. 584, 3311–3316. doi: 10.1016/j.febslet.2010.07.012
Ulke-Lemee, A., Sun, D. H., Ishida, H., Vogel, H. J., and MacDonald, J. A. (2017). Binding of smoothelin-like 1 to tropomyosin and calmodulin is mutually exclusive and regulated by phosphorylation. BMC Biochem. 18:5. doi: 10.1186/s12858-017-0080-6
Ulke-Lemee, A., Turner, S. R., Mughal, S. H., Borman, M. A., Winkfein, R. J., and MacDonald, J. A. (2011). Mapping and functional characterization of the murine smoothelin-like 1 promoter. BMC Mol. Biol. 12:10. doi: 10.1186/1471-2199-12-10
Wang, Y., Deng, W., Zhang, Y., Sun, S., Zhao, S., Chen, Y., et al. (2018). MICAL2 promotes breast cancer cell migration by maintaining epidermal growth factor receptor (EGFR) stability and EGFR/P38 signalling activation. Acta Physiol. 222, e12920. doi: 10.1111/apha.12920
Wang, Z., Li, T. E., Chen, M., Pan, J. J., and Shen, K. W. (2020). miR-106b-5p contributes to the lung metastasis of breast cancer via targeting CNN1 and regulating Rho/ROCK1 pathway. Aging 12, 1867–1887. doi: 10.18632/aging.102719
Yao, J., Wu, X., Zhang, D., Wang, L., Zhang, L., Reynolds, E. X., et al. (2019). Elevated endothelial Sox2 causes lumen disruption and cerebral arteriovenous malformations. J. Clin. Invest. 129, 3121–3133. doi: 10.1172/JCI125965
Yin, L. M., Ulloa, L., and Yang, Y. Q. (2019). Transgelin-2: biochemical and clinical implications in cancer and asthma. Trends Biochem. Sci. 44, 885–896. doi: 10.1016/j.tibs.2019.05.004
Yin, L. M., Xu, Y. D., Peng, L. L., Duan, T. T., Liu, J. Y., Xu, Z., et al. (2018). Transgelin-2 as a therapeutic target for asthmatic pulmonary resistance. Sci. Transl. Med. 10:eaam8604. doi: 10.1126/scitranslmed.aam8604
Keywords: actin cytoskeleton, CH-domain-containing proteins, α-helix, tubulin, calmodulin, tropomyosin, transgelin-2, cancer
Citation: Yin L-M, Schnoor M and Jun C-D (2020) Structural Characteristics, Binding Partners and Related Diseases of the Calponin Homology (CH) Domain. Front. Cell Dev. Biol. 8:342. doi: 10.3389/fcell.2020.00342
Received: 10 March 2020; Accepted: 20 April 2020;
Published: 14 May 2020.
Edited by:
Vladimir Sytnyk, University of New South Wales, AustraliaReviewed by:
Mark Pfuhl, King’s College London, United KingdomOrest William Blaschuk, McGill University, Canada
Copyright © 2020 Yin, Schnoor and Jun. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lei-Miao Yin, collegeylm@shutcm.edu.cn; collegeylm@163.com