 Carlos R. Magni1,2
Carlos R. Magni1,2 Paola L. Poch1,3
Paola L. Poch1,3 Sergio E. Espinoza4*
Sergio E. Espinoza4* Marco A. Yáñez1Eduardo E. Martínez1,2
Marco A. Yáñez1Eduardo E. Martínez1,2 Alvaro A. Promis2
Alvaro A. Promis2 Gabriel A. Mancilla2,5
Gabriel A. Mancilla2,5- 1Facultad de Ciencias Forestales y de la Conservación de la Naturaleza, CESAF Universidad de Chile, Santiago, Chile
- 2Departamento de Silvicultura y Conservación de la Naturaleza, Facultad de Ciencias Forestales y de la Conservación de la Naturaleza, Universidad de Chile, Santiago, Chile
- 3Instituto de Ecología y Biodiversidad (IEB), Santiago, Chile
- 4Facultad de Ciencias Agrarias y Forestales, Universidad Católica del Maule, Talca, Chile
- 5Centro del Agua para Zonas Áridas y Semiáridas de América Latina y el Caribe (CAZALAC), La Serena, Chile
The higher drought intensity recorded in the last decades in the Mediterranean zone of Chile increased the level of threats of the endemic Beilschmiedia miersii (Gay) Kosterm and has raised concern about its capacity to face drought. In this study, we assessed the effect of seed provenance and mother tree on seed germination traits, seedling growth, biomass allocation, presence of cotyledons, and survival responses of B. miersii cultivated under two water treatments (well-watered versus water stress. Average pre-dawn plant water potentials of −0.9 and −4.5 MPa, respectively). The germination stage of this trial showed significant differences in germination capacity, germination energy, and the maximum value of Czabator. Provenances from the southern range of the species, with higher precipitation (Cantillana and El Arbol) had a better germination performance, however, once germinated, the ability of the seedlings to survive in the nursery was high only in two of the five provenances under study. The coastal and interior provenances with lower precipitations (El Pobre, Longotoma, and Antumapu) were not able to survive to the seedling stage (<5% survival). Seedling growth and survival varied significantly between the two remaining provenances and the two watering treatments. The provenance El Arbol, exhibited a higher survival, growth, presence of cotyledons, and biomass traits. This provenance also exhibited a higher presence of cotyledons in both watering treatments and an unaltered root to shoot ratio between the well-watered and the water stress treatments. Our results suggests that there is important phenotypic variation for seed germination and seedling survival associated to the provenance origin, emphasizing the importance of a nursery-evaluation phase before the beginning of restoration projects at the landscape level.
1. Introduction
Seed source studies in forest trees are a valuable tool to identify better seed sources when planning ecological restoration actions, particularly in regions where the Mediterranean drylands are affected by drought (Broadhurst and Boshier, 2014). In these areas, the restoration of Mediterranean-type ecosystems has had limited success because climate warming and increased aridification are changing the habitat to which populations have been adapted, altering species range, phenology and vegetation types, and threatening tree recruitment and survival (Vallejo et al., 2012; Benito-Garzón et al., 2013; Gazol et al., 2018). Besides, factors such as anthropogenic disturbances (Mandle et al., 2011), lack of governmental restoration plans, poor quality and quantity of the planting stock, low survival after outplanting (Bannister et al., 2019; Acevedo et al., 2021), and lack of adequate seed supply (León-Lobos et al., 2020) constrain forest landscape restoration. In addition, there is lack of information on the early responses of seedlings to water stress, as a tool to guide the selection of appropriate seed sources for restoration.
In the Mediterranean-type ecosystems of central Chile Beilschmiedia miersii (Gay) Kosterm (Lauraceae) is an endemic and threathened recalcitrant-seeded tree species with hypogeal germination, a very limited distributional range (Henríquez and Simonetti, 2001) and the largest seeds among Mediterranean flora (Fuentes et al., 1986). Its overall geographical range extends no more than 200 km from coastal areas (32° S) and altitudes close to the sea level, to interior areas (34° S) with altitudes close to 1,200 m.a.s.l (Donoso, 1998). The species has been considered as Vulnerable (Benoit, 1989; MINSEGPRES, 2008) because of threats such as land conversion for agricultural uses, damage by livestock, leaf litter removal, drought (Henríquez and Simonetti, 2001; Becerra et al., 2004; Kremer et al., 2019) and fire (de la Barrera et al., 2018). Recently, Venegas-González et al. (2022) reported a decline in growth of B. miersii forests since the early 1900s as as consequence of climatic variability. Besides, seed viability has been negatively affected by rodents, insects and other herbivores, diminishing the sexual regeneration of the species (Serra et al., 1986; Gajardo et al., 1987; Henríquez and Simonetti, 2001; Morales et al., 2015).
Adaptations to ecological conditions can lead to differences in growth performance in seedlings of different provenances (Gratani et al., 2003; Vasques et al., 2013) and trees respond to changes in environmental conditions through phenotypic plasticity (i.e, the ability of a genotype to alter its morphology and physiology in response to changes in the environmental conditions) (Bradshaw, 1965; Nicotra et al., 2010), which allows species to grow in a range of environments (Corcuera et al., 2010). Beilschmiedi miersii is a relict species (Cabrera, 1959) that has been typically restricted to humid and shaded environments (Becerra et al., 2004; Novoa, 2004), but phenotypic plasticity has allowed populations to prosper under different environmental conditions in the last decades. The geographic area were the species develops has been subjected to an uninterrupted sequence of dry years from 2010 to 2018 (Garreaud et al., 2020) with the consequent negative effects on vegetation development (Garreaud et al., 2017; Miranda et al., 2020). In this context, given the contrasting environmental conditions within the limited area where B. miersii currently develops, it is expected phenotypic variation in seed germination and seedling responses, especially to environmental stresses such as drought, which is projected to increase in areas with Mediterrarean-type climate (Seager et al., 2019). However, population growth is not always related to climate variation (Peterson et al., 2021), and in B. miersii this is still poorly understood. Studies with species from Mediterranean-type climate areas (e.g., Quercus ilex L. and Quillaja saponaria Mol.) have shown that seed germination and seedling survival is high in provenances with annual precipitation close to 500 mm, whereas the lowest germination and survival values were reported in provenances with annual precipitation close to 900 mm (Caliskan, 2014; Magni et al., 2019). In field experiments with B. miersii it was found that seedling survival improved with irrigation, and the response depended on the site canopy conditions and the presence of leaf litter (Kremer et al., 2019), whereas, under laboratory conditions, seedling survival was not affected by irrigation (Becerra et al., 2004). However, as these studies only used one local provenance origin, the results are limited geographycally because it is known that performance increases as local seed is planted near to their home site (Joshi et al., 2001).
Provenance differentiation for seed germination and seedling growth in Chilean native species has been investigated at the field (Espinoza et al., 2020) and nursery level (Magni et al., 2019). Seedling performance in large- recalcitrant seed species is influenced by traits such as the root to shoot ratio (RSR) and the presence of cotyledons. The RSR confers adaptability to harsh environments because it is a critical factor for optimizing the balance between water uptake and loss to avoid summer drought damage (Grossnickle, 2000), whereas the cotyledons store nutritional substances that determine the initial development of seedlings (Yi and Wang, 2016). However, there is still little knowledge about the provenance variation and the phenotypic variation of different seed sources to stresses such as drought at the seedling stage, as a tool to guide the restoration of B. miersii. This information is relevant to select better seed sources and diminish the high mortality rate observed in seedling establishment of woody Mediterranean species (Davis, 1989). As young seedlings are more susceptible to drought than adult trees (Cavender-Bares and Bazzaz, 2000), information at the seedling stage might give indications of the capacity of a species to grow and survive under water- restricted environments. In this context, the aim of this study was to analyze the variation for seed germination and the phenotypic responses to water restriction of B. miersii provenances. We analyzed germination traits in five provenances (seed sources) with germinated seeds, and then we assessed responses in survival, growth and biomass partitioning under contrasting watering regimes in two provenances that survived to seedling stage. As B. miersii is a recalcitrant-seeded species that needs moisture to germinate, we expect a better germination capacity and seedling performance from the wettest provenances and higher RSR in seedlings from the drier provenances.
2. Material and methods
2.1. Plant material
This study included seeds from five provenance origins from coastal and inland areas of Central Chile (Table 1). These provenances comprise the narrow range of distribution (maximum distance of 190 km) and environmental conditions where the species currently develops. From February to early April 2013, ripe seeds were directly collected from trees on each provenance and the plantation from 40 mother trees randomly selected, and stored at 4°C under similar conditions until use. ISTA standards were followed to clean seeds (ISTA, 2006). The number of mother trees varied by provenance (Table 1). The study was conducted in a nursery located at the Universidad de Chile, La Pintana, Chile (33° 34′ S, 70° 37′ W, 611 m.a.s.l) and in April 2013 the seeds were soaked in tap water for 24 h, and those that floated and with insect damage were discarded. Approximately sixteen viable seeds from each of 40 mother trees (i.e., a total of 3,168 seeds) were sown in 150-mL pots filled with a mixture of composted bark, local topsoil, and sand (1:1:1 v). Then, they were cultured under similar ambient conditions during 12 months until early April 2014 under daily irrigation according to the season of the year (1.25 L m2 in fall-spring, and 5 L m2 during the summer months). No fertilizer was added to the growing substrate.
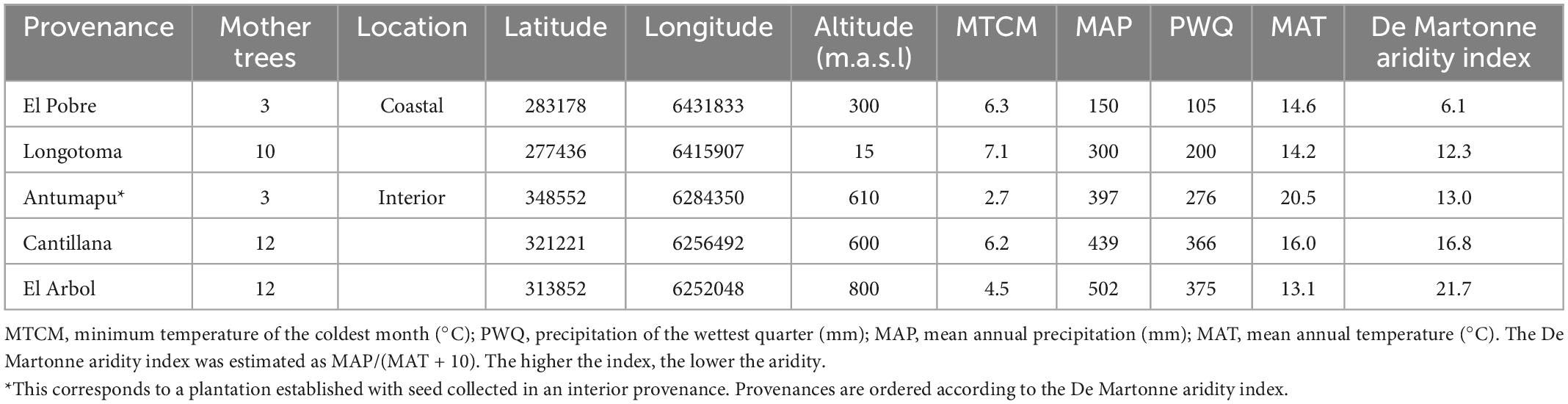
Table 1. Location and climate for the provenances and plantation of Beilschmiedi miersii under study.
2.2. Assessment of seed germination
Germination was recorded daily on those seeds with emerged cotyledons. The germination capacity (GC) was obtained as the proportion between the germinated seeds and the total seeds sown. The maximum value of Czabator (MV) was obtained as the maximum quotient derived from all of the cumulative full-seed germination percentages on any day divided by the number of days to reach these percentages (Czabator, 1962). Additionally, we determined the germinative energy (GE), as the accumulated percentage of germination on the day that MV occurs, and the energy period (EP), as the number of days in which MV occurs (FAO, 1985; ISTA, 2006).
2.3. Seedling experiment and watering treatments
After the seed germination finished the provenances El Pobre, Longotoma, and Antumapu were excluded from the analysis due to the low number of germinated seedlings. Provenances Cantillana and El Arbol were represented with 7 and 11 mother trees (synonym families and coded as C and A, respectively), and the total number of seedlings in this phase was 1,265. The watering experiment was established in April 2014 with the germinated seeds of the germination experiment (i.e., 1-year-old seedlings) and it was arranged following a split-plot in an incomplete block design with 11 replicates. Two watering treatments were the whole plot treatment [well-watered (WW) and water stress (WS)], and the provenances were the split-plot treatment. As both provenances had an heterogenous number of germinated seedlings, we used incomplete blocks but we took care that each family was present in more than three replicates in each watering treatment and containing at least three seedlings. Watering regimes were defined based on pre-dawn plant water potentials (ψpd) measured by using a Scholander pressure chamber (PMS Instrument, Albany, USA). In the WW treatment, seedlings were watered daily until reaching an average ψpd of −0.9 MPa. In the WS treatment, water was withheld from April 11th to May 6th 2014 (i.e., one 26-days cycle of water withholding) and seedlings reached an average ψpd of −4.5 MPa.
2.4. Assessments
Seedling height (H, cm), root collar diameter (D, mm), and survival (SUR, %), and the presence of cotyledons in each seedling (COT, %) were measured after the watering treatments had finished. SUR and COT were assessed as categorical variables. For SUR, 1 represented an alive seedling, and 0 represented a dead seedling. For cotyledons, 1 represented a present cotyledon, and 0 represented an absent cotyledon. Afterward, cotyledons were extracted from each seedling and weighed at ± 0.01 (Wcot, g). Finally, all seedlings were harvested and oven-dried at 65°C until constant weight. The dry weight fractions of roots (RDW), leaves (LDW), stems (SDW) were determined (±0.01, g). The total dry weight (TDW) was derived as RDW+LDW+SDW, and the root:shoot ratio (RSR) was derived as RSR = RDW/(SDW+LDW).
2.5. Data analyses
Seed germination traits were analyzed using the full data set (n = 3,168 sown seeds) of the four provenances and the plantation, and the linear mixed model presented in Equation 1. Otherwise, survival, growth, cotyledons, and biomass traits were analyzed using a reduced dataset containing only seedlings from provenances Cantillana and El Arbol (n = 1,265 seedlings), and the linear mixed model presented in Equation 2. We examined the assumptions for normality and homogeneity of variance required for the analysis of variance, and used Box-Cox transformation when neccesary. The survival and the presence of cotyledons; measured as categorical values (i.e., 1, 0), were analyzed by using a generalized linear model with binomial distribution and a logit link function (Stroup, 2012). The analyses of variance for each trait was carried out on a family-mean basis.
Where, Y is the observed phenotypic value, μ is the overall mean, W is the fixed effect of watering regime, R is the fixed effect of replicate, P is the fixed effect of provenance, M(P) is the random effect of the mother tree nested within the provenance. In Equation 1, e represents the experimental random error. In Equation 2, e1 is the error associated with the whole plot, and e2 is the error associated with the split-plot. Mean comparisons were made using the Tukey test at P ≤ 0.05. In this equation, a significant effect of W indicates the existence of phenotypic plasticity in response to water stress for the trait considered, whereas a significant genotype by environment interaction (i.e., interaction W × P), indicates the existence of genetic differences in plasticity (Schlichting, 1986).
Because our interest was also to analyze the survival of seedlings in the WS treatment, we used the reduced dataset containing two provenances (n = 1,265 seedlings) to conduct simple regression analysis with family means in order to assess the relationships between seedling SUR with seedling responses in growth (D and H), biomass partitioning (SDW, LDW, RDW, RSR, and TDW), and cotyledons (COT and Wcot). We checked assumptions of normality and homogeneity of variances and the goodness of fit for the regression model was assessed by computing the coefficient of determination and the Pearson’s coefficient of correlation. In addition, we used Path analysis to quantify simultaneously the direct and indirect contributions of seedling attributes on seedling survival. We assessed the influence of D, H, RDW, LDW, SDW, COT, and Wcot on seedling SUR in the WS treatment. In this analysis we omitted those traits derived as ratios or as the sum of individual traits (i.e., RSR and TDW), to avoid multicollinearity among the independent variables. All the analysis were carried out using the SPSS software v18 (SPSS Inc, Chicago, Illinois, USA) and INFOSTAT software version 2018 (Group Infostat, Universidad Nacional de Córdoba, Argentina).
3. Results
3.1. Provenance differences in seed germination
Provenance El Arbol exhibited the highest GC, and GE, while the other provenances did not differ among them (Table 2). On average, GC and GE were 108 and 95% higher in provenance El Arbol in comparison to the other provenances. For MV, provenance El Arbol had significantly higher values than provenance El Pobre, while the other provenances had intermediate values. No differences were observed for EP in any of the provenances under study, but provenances El Pobre and Antumapu exhibited a long EP with a low variability due to the homogeneity values for this trait among mother trees.

Table 2. Means and standar error for the seed germination parameters by provenance of Beilschmiedi miersii.
3.2. Provenance differences in growth, survival, and biomass allocation under the water restriction
Before imposing the water restriction treatment, D and H were 6 and 11% higher in the provenance El Arbol than Cantillana, respectively. The magnitude of these differences between provenances was maintained after the watering restriction cycle (8 and 12%, respectively). The effect of provenance was significant for growth, biomass, survival, and cotyledons. The provenance El Arbol exhibited the highest D, H, SUR, SDW, LDW, RDW, TDW, COT, and Wcot (Table 3). The phenotypic plasticity due to the watering treatment was significant for most of the seedling traits except SDW, LDW, and Wcot, whereas the genetic differences among provenances within watering treatments (i.e., genotype by environment interaction) were significant for RSR and COT. The Provenance Cantillana had a high RSR in the WW treatment, but this was reduced in the WS treatment, while RSR in the provenance El Arbol was similar in both treatments. Moreover, RSR in Cantillana was higher than El Arbol in the WW treatment but lower in the WS treatment (Figure 1A). The provenance El Arbol had a higher percentage of cotyledons (COT) than Cantillana in both watering treatments (Figure 1B), but in both provenances COT was significantly reduced by the WS treatment. We also reported a family variation for H, SUR, COT, Wcot, RDW, and TDW (Table 3 and Figure 2), which was mainly explained by family A_16 from provenance El Arbol that exhibited the highest H, SUR, COT, RDW, and TDW, and family C_2 from provenace Cantillana, which showed the opposite pattern in almost all the abovementioned traits.
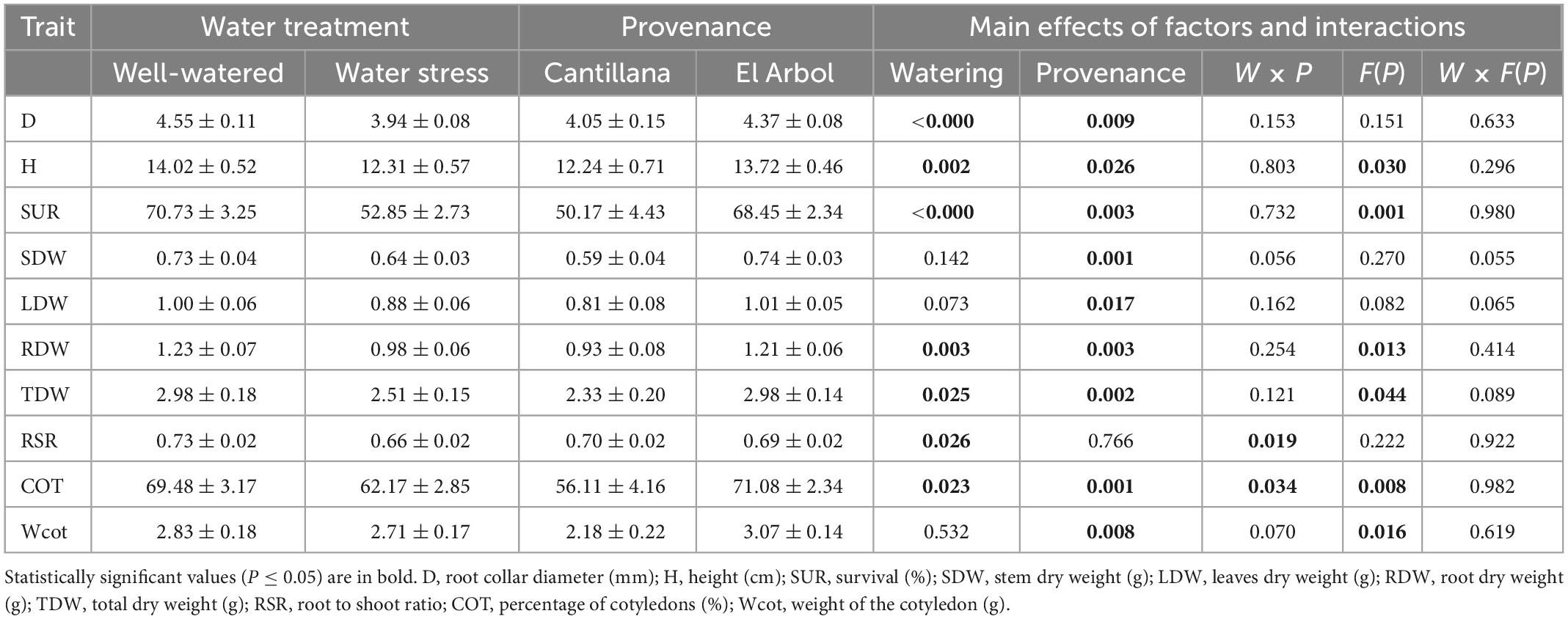
Table 3. Main effects in growth, survival, and biomass parameters tested through 2-Way ANOVA using watering treatment (W) and provenance (P) as fixed factors and family (F) as a random nested factor in provenance (F(P)).
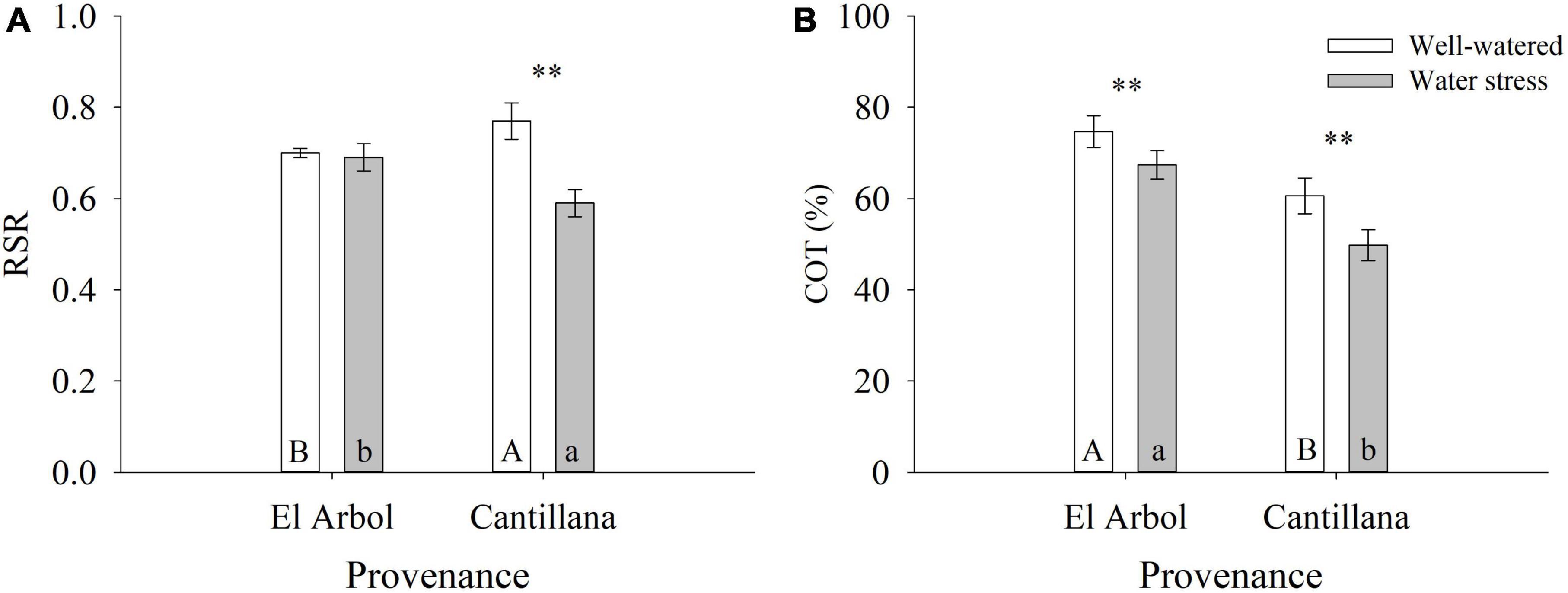
Figure 1. Provenance variation by watering regime for RSR (A) and COT (B). **Indicate significant differences within a provenance at P ≤ 0.05. The same lowercase and uppercase letters indicate no significant differences (P ≤ 0.05) between the provenances in the well-watered and water stress treatment.
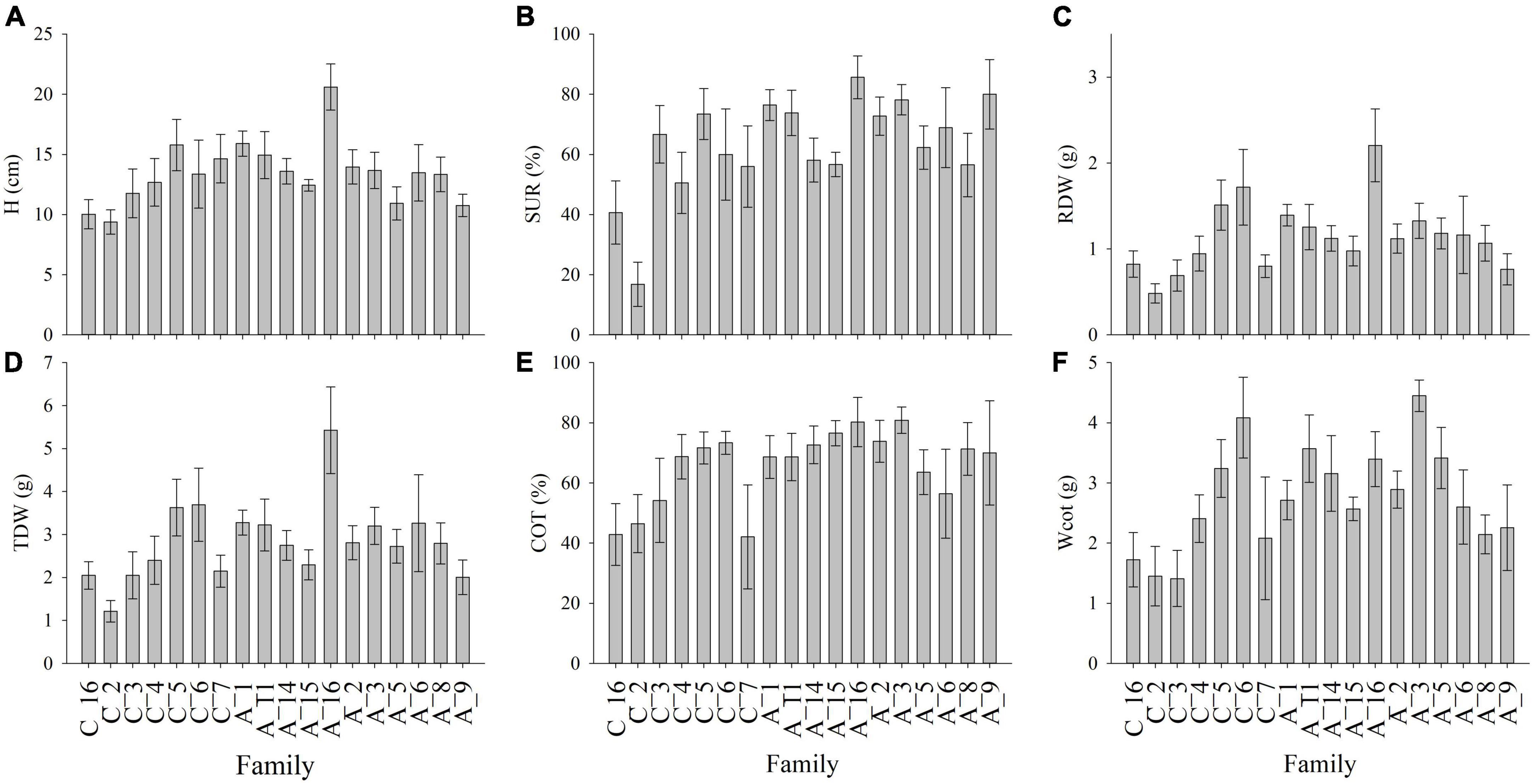
Figure 2. Family variation for growth, survival, biomass allocation and weight of cotyledons in seedlings of Beilschmiedia miersii submitted to contrasting watering regimes. (A) H, height; (B) SUR, survival; (C) RDW, roots dry weight; (D) TDW, total dry weight; (E) COT, percentage of cotyledons; and (F) Wcot, weight of the cotyledon. C, families from provenance Cantillana; A, families from provenance El Arbol. Numbers after the dash corresponds to family number.
3.3. Correlation between seed and seedling characteristics
When comparing the correlations among survival with seedling responses in growth, biomass and cotyledons characteristics; with the exception of RSR, they were mostly medium-low and positive correlations (Figure 3). However, the Path analysis indicated that only H and COT had significant direct and positive effects on seedling survival (Table 4). We also observed positive and indirect effects of the other traits under study. The trait with the greatest indirect effect on seedling SUR was D, followed by LDW, RDW, Wcot and SDW. Consistently, those trats, via positively affecting H, indirectly affected seedling SUR. In the case of Wcot, via positively affecting COT, indirectly affected seedling SUR.
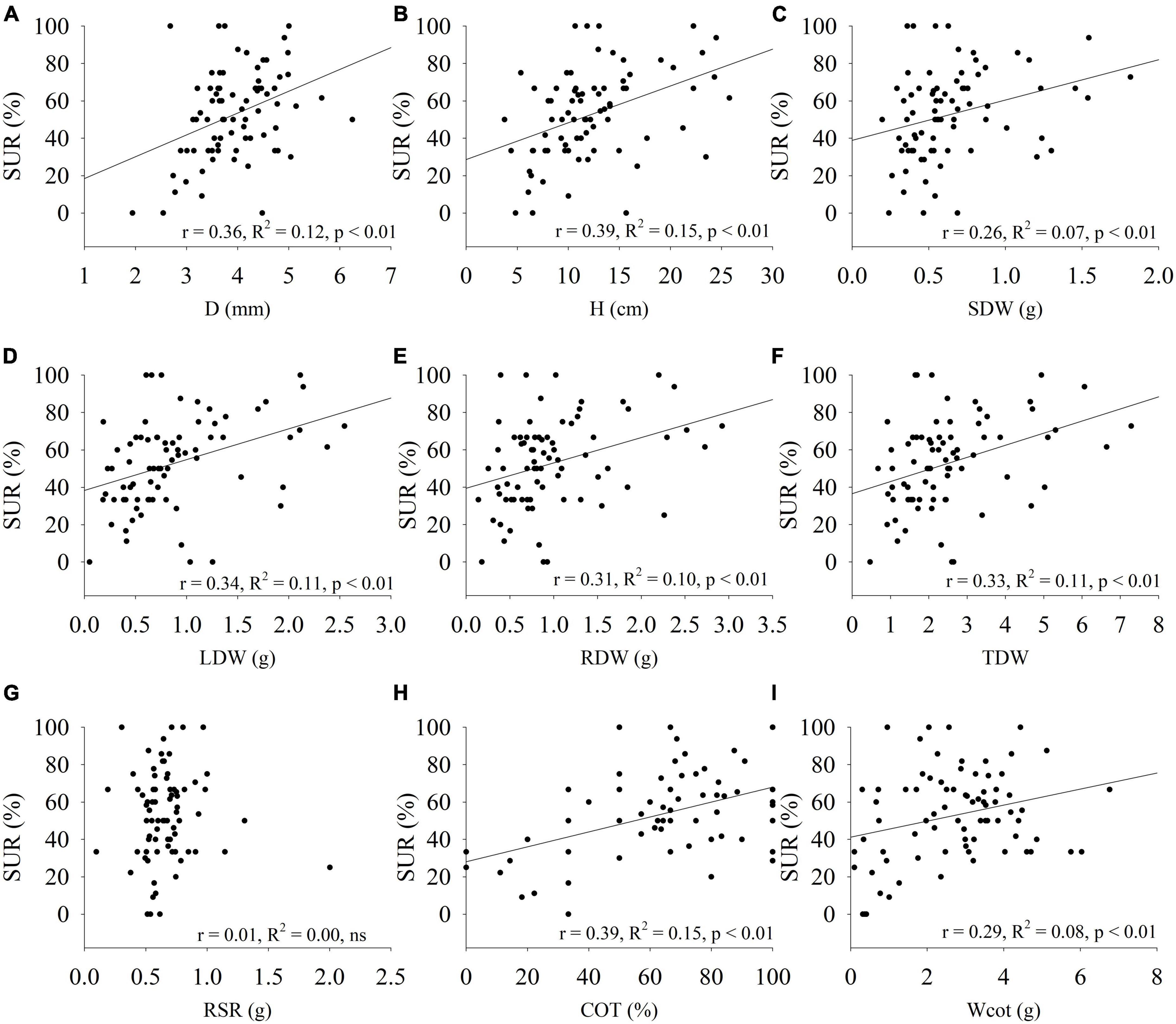
Figure 3. Relationship among the seedling survival with seedling traits of Beilschmiedi miersii in the WS treatment in provenances Cantillana and El Arbol. Regressions were conducted with average family values in a replicate. (A) D, root collar diameter; (B) H, height; (C) SDW, stem dry weight; (D) LDW, leaves dry weight; (E) RDW, root dry weight; (F) TDW, total dry weight; (G) RSR, root to shoot ratio; (H) COT, percentage of cotyledons; and (I) Wcot, weight of the cotyledon.
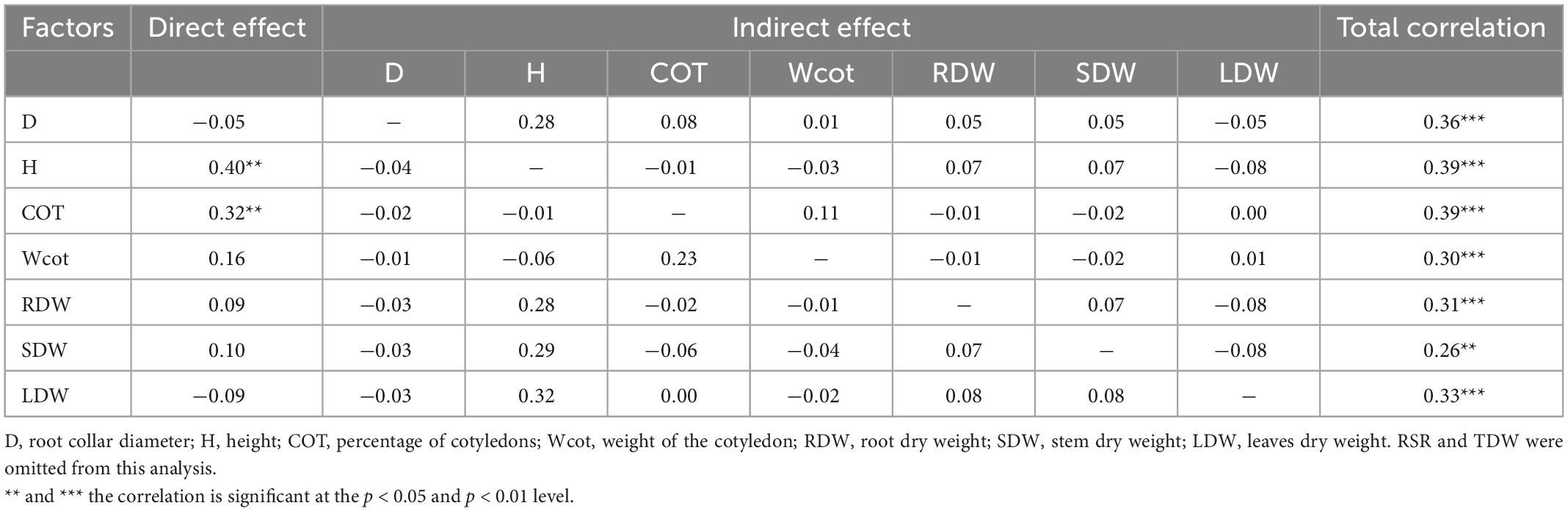
Table 4. Path analysis of the relationships between seedling survival in the WS treatment and the various independent variables (growth, biomass and cotyledons).
4. Discussion
4.1. Provenance differences in germination traits
Our results indicated that the provenance effect considerably impacted the germination traits. GC, GE, and MV of B. miersii were significantly higher in the El Arbol provenance seed and a large number of seeds were able to survive to the seedling stage from this provenance. The coastal provenances El Pobre and Longotoma and the interior plantation Antumapu were not able to survive in large number to the seedling stage, which might be because those provenances are located in the dry edge of the species distribution (i.e., the lower the De Martonne aridity index in Table 1), with the consequent low individual performance (Brown, 1984). This lack of ability to survive to the seedling stage might be also related to lack of pollinators and inbreeding in those populations. A worldwide decline of pollinators (Rodger et al., 2021) has critically impacted the Mediterranean-type ecosystem of Chile (Vieli et al., 2021).
The differences in precipitation, altitude, and temperature across provenances, might have also caused the variations in germination capacity. B. miersii is a recalcitrant seeded-species that needs moisture to germinate. Recalcitrant seeds are highly sensitive to desiccation and lose their viability in dry sites (Bewley et al., 2006). The cumulated precipitation from the year before the seed release to the month of seed dispersal and seed collection (i.e., from January 2012 to April 2013) averaged 212 mm in the three coastal provenances, while the interior provenance El Arbol averaged 529 mm of precipitation in the same period.1 In this context, the higher precipitation in El Arbol than in the other provenances may partially explain the differences in seed germination, but this needs further research because we used only viable seed in our experiment.
On the other hand, provenances that survived to the seedling stage (Cantillana and El Arbol) come from areas with higher altitude (close to 800 m.a.s.l.), while those provenances that were not able to survive to the seedling stage are mainly from coastal areas close to the sea level. Provenances Longotoma and El Arbol have similar annual temperature but they differed in altitude and aridity, with the latter provenance being present at higher altitudes with higher precipitation and lower aridity. In general, seeds from higher altitudes have higher germination rate than those from the lower altitudes, which is related to the size of cotyledons, i.e., seed from higher altitudes are heavier than those of lower altitudes because they have to survive under severe environments (Mariko et al., 1993). Average winter temperature in provenances El Arbol is 2.8°C, compared to the 6.9°C of coastal provenances (see text footnote 1).
Due to the large number (3,168) and size of seeds, we assessed viability according to the simple floating method and not by more accurate methods (i.e., cutting and tetrazolium methods). Thus, another explanation for the lower germination rate experienced by coastal provenances might be the presence of insect-infested seeds; probably by Hypothenemus sp. (data not shown), that did not float in the test. The acorns of large-seeded species infested by insects experience lower germination (Branco et al., 2002; Yi and Zhang, 2008) and B. miersii has large seeds that are usually damaged during seed dispersal (Serra et al., 1986; Gajardo et al., 1987; Henríquez and Simonetti, 2001). It has been reported that damaged seeds have a decrease in germination rate, and the surviving seedlings have low performance and high mortality (Hou et al., 2010). By visual inspection in our field campaigns, we observed low seedling regeneration in El Pobre and Longotoma provenances, but not in the Cantillana and El Arbol provenances. This could indicate a superior seedling recruitment in the latter provenances which may be associated with fewer pest insects. However, this assertion needs further research.
4.2. Provenance differences in growth, survival and biomass allocation under water restriction
In general, we found that water stress did not affect growth and survival, but affected belowground adaptations (RSR) and percentage of cotyledons (COT). Both parameters are associated with water absorption and carbohydrates reserves within the plant (Bewley and Black, 1994; Grossnickle, 2005) and may counteract decreases in carbon assimilation during water stress. As the provenances from drier environments did not survive to the seedlings stage, no assessments of RSR were made to test our hypothesis about this trait. However, the plasticity for RSR was higher in the provenance Cantillana whereas the provenance El Arbol exhibited null plasticity for this trait as it remained unaffected between watering treatments. The phenotypic plasticity of both provenances does not fit the theory of specialization proposed by Lortie and Aarssen (1996). This theory proposes that genotypes adapted to unfavorable conditions (provenance Cantillana with higher aridity), may have superior performance in restrictive environments but are unable to take advantage of favorable conditions. On the contrary, genotypes adapted to favorable environments (provenance El Arbol with lower aridity), present a superior performance in these environments, but this is reduced when conditions are limiting. According to the De Martonne index of Table 1, both provenances come from similar dry conditions (semiarid and Mediterranean type climate for Cantillana and El Arbol, respectively), thus their behavior might be explained because some organisms; particularly in the early stages of development, have the ability to alter their patterns of carbon allocation in response to certain environmental stimuli (Bloom et al., 1985). It might be possible that the low-water treatment was not sufficiently low to trigger further physiological responses in provenance El Arbol or that the presence of heavier cotyledons buffered the response. The low plasticity of El Arbol suggests a higher phenotypic stability in balancing the extraction and losses of water even in a water restricted condition, which could be linked to a specialization to adverse environments, and a conservative resource-use strategy (Baquedano et al., 2008). Seedlings with an adequate size (Villar-Salvador et al., 2012) and a balance between the shoot transpirational surface to the root absorbing surface (i.e., RSR) could have a better chance to survive in sites with water restrictions. In these sites, a high proportion of roots may ensure water supply for the plant over the first summer (Richards and Lamont, 1996). Provenance El Arbol also produced larger seedlings (H) with the highest body mass (TDW) compared with Cantillana provenance. The higher aboveground growth of the provenance El Arbol could be coupled with a higher water demands, but this is likely compensated by a greater water uptake and food reserves due to its higher mass of cotyledons (Wcot) and roots mass (RDW) in comparison to the Cantillana provenace. It is known that partial or total removal of cotyledons can reduce biomass production and even cause death in young seedlings (Armstrong and Westoby, 1993). The presence of heavier cotyledons is an adaptive character to environmental stresses (Jurado and Westoby, 1992) and might explain the high number of seeds that survive to the seedling stage in these provenances. At the mother tree level, and with the exception of mother C_6 from provenance Cantillana, most of mother trees from El Arbol provenance exhibited higher percentages and mass of cotyledons than those from Cantillana provenance. This suggests that seedling growth in B. miersii has an important maternal effect that needs to be further investigated.
4.3. Relationships between seedling survival in the WS treatment and the various independent variables
Our Path analysis provides evidence that seedling height (H) positively influenced the survival of seedlings in the water restricted treatment. The seedling diameter (D) did not directly contribute to seedling survival but did indirectly influence this trait through its positive effects on H. Height and diameter are considered good predictors of seedling survival (Grossnickle and MacDonald, 2018), and large seedlings have a better survival because of the larger roots that allow seedlings to reach moisture in deep soil horizons (Oliet et al., 2009; Cuesta et al., 2010; Villar-Salvador et al., 2012); however, larger seedlings can also have large transpiratory demands (Grossnickle, 2012) with the consequent risk of desiccation. It is known that the larger roots systems allows the supply of enough water to the foliage (Grossnickle, 2005; Manzoni et al., 2015) and the presence of heavier cotyledons promotes seedling survival because of their greater resource storage capacity (Metz et al., 2010). In our experiment, despite seedlings of provenance El Arbol were taller and had higher transpiratory surface (LDW); probably depleting the water pot more quickly than the provenance Cantillana, they had higher mass of cotyledons and root mass (Wcot, RDW). Both traits were significantly positively correlated with seedling SUR and had an indirect influence on seedling SUR via their positive effects on H and COT. Seed mass; which is a functional trait that positively affects germination and seedling survival in recalcitrant seeded species (Chacón and Bustamante, 2001), has been found to be highly correlated with storage-type cotyledons in larger seeded species (Green and Juniper, 2004). Thus, the higher Wcot and RDW might have helped to compensate larger transpiratory demands with water uptake and food reserves.
From a conservation point of view, knowledge of seed germination ability and seedling survival of B. miersii is important and may provide an insight on survival and distribution range of the species in the era of climate change. This study showed that B. miersii has phenotypic variation among provenances on seed and seedling characteristics and this variation seems to be related to the climate of the seed source origins. Provenances from driest locations had low germination capacity and were unable to survive to the seedling stage. As Mediterranean-type climate areas are expected to became drier in the coming decades (Seager et al., 2019; Fernádez et al., 2021), the natural regeneration of these provenances could be seriously compromised by the lack of water to germinate and to assure seedling establishment. Recalcitrant-seeded tree species such as B. miersii are highly sensitive to dehydration (Berjak and Pammenter, 2004), and, unlike orthodox seeds, they must germinate quickly before dehydration or die (Berjak and Pammenter, 2008). On the contrary, the provenances from sites with lower aridity produced larger seedlings with abundant and heavier cotyledons. In a global change context, this differentiation in important functional traits such as the weight of cotyledons might suggest that the southern provenance El Arbol (with heavier cotyledons to survive under harsh conditions) has the potential to provide options for re-afforestation. However, this potential needs to established in the context of future work examining fruit traits and the effect of local environmental factors on fruit development. Germinated seeds of this provenance were able to survive to the seedling stage in a severe water restricted treatment (-4.5 MPa of water potential) and were able to optimize the balance between the moisture extraction capacity of the roots relative to the transpiring shoots (RSR). In other large- recalcitrant seed species from Mediterranean-type climate areas (e.g., Quercus spp.) seedling growth and survival are negatively affected when the cotyledons are removed (Yi et al., 2019), corroborating the role of cotyledons in supporting seedling development (Yi and Wang, 2016).
Data availability statement
The data analyzed in this study is subject to the following licenses/restrictions: The data that support the findings of this study are available from the Universidad de Chile, but restrictions apply to the availability of these data, which were used under license for the current study, and so are not publicly available. Data are however available from the authors upon reasonable request and with permission of Universidad de Chile. Requests to access these datasets should be directed to CM, Y3JtYWduaUB1Y2hpbGUuY2w=.
Author contributions
CM, GM, and PP conceived and designed the study. PP performed the experiments and collected the results. SE and MY analyzed and interpreted the data, and were major contributors in writing the manuscript. AP, EM, GM, PP, CM, SE, and MY revised the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by the Fondo de Investigación del Bosque Nativo, from the Corporación Nacional Forestal (CONAF) through the project “Permanencia de Bosques de Beilschmiedia miersii. Regeneración según contenido de humedad del suelo y presencia de hojarasca” (grant number 033/2012).
Acknowledgments
We thank the staff from the Centro Productor de Semillas y Arboles Forestales (CESAF) from the Universidad de Chile, specially to Betsabé Abarca and Iván Grez.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
References
Acevedo, M., Álvarez-Maldini, C., Dumroese, R., Bannister, J., Cartes, E., and González, M. (2021). Native plant production in Chile. Is it possible to achieve restoration goals by 2035? Land 10:71. doi: 10.3390/land10010071
Armstrong, D., and Westoby, M. (1993). Seedlings from large seeds tolerate defoliation better: A test using phylogenetically in- dependent contrasts. Ecology 74, 1092–1100. doi: 10.2307/1940479
Bannister, J., Vargas-Gaete, R., Ovalle, J., Acevedo, A., Fuentes-Ramirez, A., Donoso, P., et al. (2019). Major bottlenecks for the restoration of natural forests in Chile. Restor. Ecol. 26, 1039–1104. doi: 10.1111/rec.12880
Baquedano, F., Valladares, F., and Castillo, F. (2008). Phenotypic plasticity blurs ecotypic divergence in the response of Quercus coccifera and Pinus halepensis to water stress. Eur. J. For. Res. 127, 495–506. doi: 10.1007/s10342-008-0232-8
Becerra, P., Celis-Diez, J., and Bustamante, R. (2004). Effects of leaf litter and precipitation on germination and seedling survival of the endangered tree Beilschmiedia miersii. Appl. Veg. Sci. 7, 253–257. doi: 10.1111/j.1654-109X.2004.tb00617.x
Benoit, I. (1989). Libro rojo de la flora terrestre Chilena. Santiago: Corporación Nacional Forestal.
Benito-Garzón, M., Ha-Duong, M., Frascaria-Lacoste, N., and Fernández-Manjarrés, J. (2013). Habitat restoration and climate change: Dealing with climate variability, incomplete data, and management decisions with tree translocations. Restor. Ecol. 21, 530–536. doi: 10.1111/rec.12032
Berjak, P., and Pammenter, N. (2004). “Recalcitrant seeds,” in Handbook of seed physiology: Applications to agriculture, eds R. Benech-Arnold and R. Sánchez (New York, NY: Haworth Press), 305–345.
Berjak, P., and Pammenter, N. (2008). From avicennia to zizania: Seed recalcitrance in perspective. Ann. Bot. 101, 213–228. doi: 10.1093/aob/mcm168
Bewley, J., Black, M., and Halmer, P. (2006). The encyclopedia of seeds, science technology & uses. London: CABI International.
Bewley, J., and Black, M. (1994). Seeds. Physiology of development and germination. New York, NY: Plenum Press. doi: 10.1007/978-1-4899-1002-8
Bloom, A., Chapin, F., and Mooney, H. (1985). Resource limitation in plants – an economic analogy. Ann. Rev. Ecol. Syst. 16, 363–392. doi: 10.1146/annurev.es.16.110185.002051
Bradshaw, A. (1965). Evolutionary significance of phenotypic plasticity in plants. Adv. Genet. 13, 115–155. doi: 10.1016/S0065-2660(08)60048-6
Branco, M., Branco, C., Merouani, H., and Almeida, M. (2002). Germination success, survival and seedling vigour of Quercus suber acorns in relation to insect damage. For. Ecol. Manag. 166, 159–164. doi: 10.1016/S0378-1127(01)00669-7
Broadhurst, L., and Boshier, D. (2014). “Seed provenance for restoration and management: conserving evolutionary potential and utility,” in Genetic considerartions in ecosystem restoration using native tree species, eds M. Bozzano, R. Jalonen, E. Thomas, D. Boshier, L. Gallo, S. Cavers, et al. (Rome: Food and Agriculture Organization of the United Nations), 27–38.
Brown, J. (1984). On the relationship between abundance and distribution of species. Am. Nat. 124, 255–279. doi: 10.1086/284267
Caliskan, S. (2014). Germination and seedling growth of holm oak (Quercus ilex L.): Effects of provenance, temperature, and radicle pruning. iForest 7, 103–109. doi: 10.3832/ifor0967-007
Cavender-Bares, J., and Bazzaz, F. (2000). Changes in drought response strategies with ontogeny in Quercus rubra: Implications for scaling from seedlings to mature trees. Oecologia 124, 8–18. doi: 10.1007/PL00008865
Corcuera, L., Gil-Pelegrin, E., and Notivol, E. (2010). Phenotypic plasticity in Pinus pinaster δ13C: environment modulates genetic variation. Ann For Sci. 67, 812. doi: 10.1051/forest/2010048
Cuesta, B., Villar-Salvador, P., Puértolas, J., Jacobs, D., and Rey Benayas, J. (2010). Why do large, nitrogen rich seedlings better resist stressful transplanting conditions? A physiological analysis in two functionally contrasting Mediterranean forest species. For. Ecol. Manag. 260, 71–78. doi: 10.1016/j.foreco.2010.04.002
Czabator, F. (1962). Germination value: An index combining speed and completeness of pine seed germination. For. Sci. 8, 386–396.
Chacón, P., and Bustamante, R. (2001). The effects of seed size and pericarp on seedling recruitment and biomass in Cryptocarya alba (Lauraceae) under two contrasting moisture regimes. Plant Ecol. 152, 137–144. doi: 10.1023/A:1011463127918
Davis, S. (1989). “Patterns in mixed chaparral stands: differential water status and seedling survival during summer drought,” in The California chaparral: Paradigms reexamined, ed. S. Keeley (Los Angeles, CA: Natural History Museum of Los Angeles County), 96–105.
de la Barrera, F., Barraza, P., Favier, P., Ruiz, V., and Quense, J. (2018). Megafires in Chile 2017: monitoring multiscale environmental impacts of burned ecosystems. Sci. Total Environ. 637–638, 1526–1536. doi: 10.1016/j.scitotenv.2018.05.119
Donoso, C. (1998). Bosques templados de Chile y Argentina. Variación, estructura y dinámica. Santiago: Editorial Universitaria.
Espinoza, S., Yañez, M., Magni, C., Santelices, R., and Cabrera, A. (2020). Outplanting performance of three provenances of Quillaja saponaria Mol. established in a Mediterranean drought-prone site and grown in different container size. iForest 13, 33–40. doi: 10.3832/ifor3142-012
Fernádez, A., Schumacher, V., Ciocca, I., Rifo, A., Muñoz, A., and Justino, F. (2021). Validation of a 9-km WRF dynamical downscaling of temperature and precipitation for the period 1980–2005 over Central South Chile. Theor. Appl. Climatol. 143, 361–378. doi: 10.1007/s00704-020-03416-9
Fuentes, R., Hoffmann, A., Poiani, A., and Alliende, C. (1986). Vegetation change in large clearings: patterns in Chilean matorral. Oecologia 68, 358–366. doi: 10.1007/BF01036739
Gajardo, R., Serra, M., and Grez, I. (1987). “Fichas técnicas de lugares específicos con presencia de especies leñosas amenazadas de extinción,” in Programa de protección y recuperación de la flora nativa de Chile, (Santiago: Corporación Nacional Forestal).
Garreaud, R., Alvarez-Garreton, C., Barichivich, J., Boisier, J., Christie, D., Galleguillos, M., et al. (2017). The 2010–2015 megadrought in central Chile: Impacts on regional hydroclimate and vegetation. Hydrol. Earth Syst. Sci. 21, 6307–6327. doi: 10.5194/hess-21-6307-2017
Garreaud, R., Boisier, J., Rondanelli, R., Montecinos, A., Sepúlveda, H., and Veloso-Aguila, D. (2020). The central Chile mega drought (2010–2018): A climate dynamics perspective. Int. J. Clim. 40, 421–439. doi: 10.1002/joc.6219
Gazol, A., Camarero, J., Vicente-Serrano, S., Sánchez-Salguero, R., Gutiérrez, E., de Luis, M., et al. (2018). Forest resilience to drought varies across biomes. Glob. Change Biol. 24, 2143–2158. doi: 10.1111/gcb.14082
Gratani, L., Meneghini, M., Pesoli, P., and Crescente, M. (2003). Structural and functional plasticity of Quercus ilex seedlings of different provenances in Italy. Trees 17, 515–521. doi: 10.1007/s00468-003-0269-8
Green, P., and Juniper, P. (2004). Seed–seedling allometry in tropical rain forest trees: Seed mass-related patterns of resource allocation and the “reserve effect”. J. Ecol. 92, 397–408. doi: 10.1111/j.0022-0477.2004.00889.x
Grossnickle, S. (2000). Ecophysiology of northern spruce species: The performance of planted seedlings. Ottawa: NRC Press.
Grossnickle, S. (2005). Importance of root growth in overcoming planting stress. New For. 30, 273–294. doi: 10.1007/s11056-004-8303-2
Grossnickle, S., and MacDonald, J. (2018). Why seedlings grow: Influence of plant attributes. New For. 49, 1–34. doi: 10.1007/s11056-017-9606-4
Grossnickle, S. (2012). Why seedlings survive: Influence of plant attributes. New For. 43, 711–738. doi: 10.1007/s11056-012-9336-6
Henríquez, C., and Simonetti, J. (2001). The effect of introduced herbivores upon an endangered tree (Beilschmiedia miersii. Lauraceae). Biol. Cons. 98, 69–76. doi: 10.1016/S0006-3207(00)00143-9
Hou, X., Yi, X., Yang, Y., and Liu, W. (2010). Acorn germination and seedling survival of Q. variabilis: Effects of cotyledon excision. Ann. For. Sci. 67, 711. doi: 10.1051/forest/2010036
ISTA (2006). International rules for seed testing. Edition 2006. Bassersdorf: International Seed Testing Association (ISTA).
Joshi, J., Schmid, B., Caldeira, M., Dimitrakopoulos, P., Good, J., Harris, R., et al. (2001). Local adaptation enhances performance of common plant species. Ecol. Lett. 4, 536–544. doi: 10.1046/j.1461-0248.2001.00262.x
Jurado, E., and Westoby, M. (1992). Seedling growth in relation to seed size among species of arid Australia. J. Ecol. 80, 407–416. doi: 10.2307/2260686
Kremer, K., Promis, ÁA., Mancilla, G., and Magni, C. (2019). Leaf litter and irrigation can increase seed germination and early seedling survival of the recalcitrant-seeded tree Beilschmiedia miersii. Austral Ecol. 44, 86–94. doi: 10.1111/aec.12655
León-Lobos, P., Bustamente-Sánchez, M., Nelson, C., Alarcón, D., Hasbún, R., Way, M., et al. (2020). Lack of adequate seed supply is a major bottleneck for effective ecosystem restoration in Chile: Friendly amendment to Bannister et al. (2018). Restor. Ecol. 28, 277–281. doi: 10.1111/rec.13113
Lortie, C., and Aarssen, L. (1996). The specialization hypothesis for phenotypic plasticity in plants. Int. J. Plant Sci. 157, 484–487. doi: 10.1086/297365
Magni, C., Espinoza, S., Poch, P., Abarca, B., Grez, I., Martínez, E., et al. (2019). Growth and biomass partitioning of nine provenances of Quillaja saponaria seedlings to water stress. South For. 81, 103–109. doi: 10.2989/20702620.2018.1512789
Manzoni, S., Vico, G., Thompson, S., Beyer, F., and Weih, M. (2015). Contrasting leaf phenological strategies optimize carbon gain under droughts of different duration. Adv. Water Resour. 84, 37–51. doi: 10.1016/j.advwatres.2015.08.001
Mandle, L., Bufford, J., Schmidt, I., and Daehler, C. (2011). Woody exotic plant invasions and fire: Reciprocal impacts and consequences for native ecosystems. Biol. Invasions 13, 1815–1827. doi: 10.1007/s10530-011-0001-3
Mariko, S., Koizumi, H., Suzuki, J., and Furukawa, A. (1993). Altitudinal variations in germination and growth responses of Reynoutria japonica populations on Mt. Fuji to a controlled thermal environment. Ecol. Res. 8, 27–34. doi: 10.1007/BF02348604
Metz, J., Liancourt, P., Kigel, J., Harel, D., Sternberg, M., and Tielbörger, K. (2010). Plant survival in relation to seed size along environmental gradients: A long-term study from semi-arid and Mediterranean annual plant communities. J. Ecol. 98, 697–704. doi: 10.1111/j.1365-2745.2010.01652.x
MINSEGPRES (2008). (Ministerio Secretaría General de la Presidencia, CL). Decreto Supremo No 50/2008. Chile. Aprueba y oficializa nómina para el segundo proceso de clasificación de especies según su estado de conservación. Available online at: https://www.bcn.cl/leychile/navegar?idNorma=273234&idParte=0 (accessed on March 25, 2021).
Miranda, A., Lara, A., Altamirano, A., Di Bella, C., González, M., and Camarero, J. (2020). Forest browning trends in response to drought in a highly threatened mediterranean landscape of South America. Ecol. Indic. 115:106401. doi: 10.1016/j.ecolind.2020.106401
Morales, N., Becerra, P., Arellano, E., and Gilabert, H. (2015). Effect of large and small herbivores on seed and seedling survival of Beilschmiedia miersii in central Chile. Bosque 36, 127–132. doi: 10.4067/S0717-92002015000100014
Nicotra, A., Atkin, O., Bonser, S., Davidson, A., Finnegan, E., Mathesius, U., et al. (2010). Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 15, 684–692. doi: 10.1016/j.tplants.2010.09.008
Novoa, P. (2004). Determinación del grado de amenaza del belloto del norte (Beilschmiedia miersii Kosterm, Lauraceae), mediante el uso de la metodología UICN 2001 (Version 3.1). Chloris Chilensis. 7.
Oliet, J., Planelles, Z., Artero, Z., Valverde, Z., Jacobs, D., and Segura, Z. (2009). Field performance of Pinus halepensis planted in Mediterranean arid conditions: Relative influence of seedling morphology and mineral nutrition. New For. 37, 313–331. doi: 10.1007/s11056-008-9126-3
Peterson, M., Bailes, G., Hendricks, L., Pfeifer-Meister, L., Reed, P., Bridgham, S., et al. (2021). Latitudinal gradients in population growth do not reflect demographic responses to climate. Ecology 31:e02242.1.
Richards, M., and Lamont, B. (1996). Post-fire mortality and water relations of three congeneric shrub species under extreme water stress-a trade-off with fecundity? Oecologia 107, 53–60. doi: 10.1007/BF00582234
Rodger, J., Bennett, J., Razanajatovo, M., Knight, T., van Kleunen, M., Ashman, T., et al. (2021). Widespread vulnerability of flowering plants seed production to pollinator declines. Sci. Adv. 7:eabd3524. doi: 10.1126/sciadv.abd3524
Seager, R., Osborn, T., Kushnir, Y., Simpson, I., Nakamura, J., and Liu, H. (2019). Climate variability and change of Mediterranean-type climates. J. Clim. 32, 2887–2915. doi: 10.1175/JCLI-D-18-0472.1
Serra, M., Gajardo, R., and Cabello, A. (1986). Beilschmiedia miersii (Gay) Kosterm. “Belloto” (Lauraceae). Ficha teécnica de especies amenazadas. Programa de protección y recuperacioón de la flora nativa de Chile. Santiago: Corporación Nacional Forestal.
Schlichting, C. (1986). The evolution of phenotypic plasticity in plants. Ann. Rev. Ecol. Syst. 17, 667–693. doi: 10.1146/annurev.es.17.110186.003315
Stroup, W. (2012). Generalized Linear Mixed Models: Modern Concepts, Methods, and Applications. Boca Raton: CRC Press.
Vallejo, R., Smanis, A., Chirino, E., Fuentes, D., Valdecantos, A., and Vilagrosa, A. (2012). Perspectives in dryland restoration: Approaches for climate change adaptation. New For. 43, 561–579. doi: 10.1007/s11056-012-9325-9
Vasques, A., Chirino, E., Vilagrosa, A., Vallejo, V., and Keizer, J. (2013). The role of seed provenance in the early development of Arbutus unedo seedlings under contrasting watering conditions. Environ. Exp. Bot. 96, 11–19. doi: 10.1016/j.envexpbot.2013.08.004
Venegas-Gonzá,lez, A., Muñoz, A., Carpintero-Gibson, S., González-Reyes, A., Schneider, I., Gipolou-Zuñiga, T., et al. (2022). Sclerophyllous forest tree growth under the influence of a historic megadrought in the mediterranean ecoregion of Chile. Ecosystems 1–18. doi: 10.1007/s10021-022-00760-x
Vieli, L., Murúa, M., Flores-Prado, L., Carvallo, G., Valdivia, C., Muschett, G., et al. (2021). Local actions to tackle a global problem: A multidimientional assessment of the pollination crisis in Chile. Diversity 13, 2–18. doi: 10.3390/d13110571
Villar-Salvador, P., Puértolas, J., Cuesta, B., Peñuelas, J., Uscola, M., Heredia-Guerrero, N., et al. (2012). Increase in size and nitrogen concentration enhances seedling survival in Mediterranean plantations. Insights from an ecophysiological conceptual model of plant survival. New For. 43, 755–770. doi: 10.1007/s11056-012-9328-6
Yi, X., and Zhang, Z. (2008). Influence of insect-infested cotyledons on early seedling growth of Mongolian oak, Quercus mongolica. Photosynthetica 46, 139–142. doi: 10.1007/s11099-008-0022-z
Yi, X., Bartlow, A., Curtis, R., Agosta, S., and Steele, M. (2019). Responses of seedling growth and survival to post-germination cotyledon removal: An investigation among seven oak species. J. Ecol. 107, 1817–1827. doi: 10.1111/1365-2745.13153
Keywords: provenance origin, water restriction, restricted range species, Mediterranean-type ecosystems, recruitment, restoration, recalcitrant-seeded tree species
Citation: Magni CR, Poch PL, Espinoza SE, Yáñez MA, Martínez EE, Promis AA and Mancilla GA (2023) Provenance influences seed germination and phenotypic responses to water restriction in the endemic Beilschmiedia miersii (Gay) Kosterm. Front. For. Glob. Change 5:1039908. doi: 10.3389/ffgc.2022.1039908
Received: 08 September 2022; Accepted: 30 December 2022;
Published: 19 January 2023.
Edited by:
Narkis Morales, Pontificia Universidad Católica de Chile, ChileReviewed by:
Jose Climent, Instituto Nacional de Investigación y Tecnología Agroalimentaria (INIA), SpainManuel Esperon-Rodriguez, Western Sydney University, Australia
Copyright © 2023 Magni, Poch, Espinoza, Yáñez, Martínez, Promis and Mancilla. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sergio E. Espinoza,  ZXNwaW5vemFAdWNtLmNs
ZXNwaW5vemFAdWNtLmNs